2025-07-31 阅读量:1722
在神经环路研究领域,睡眠、内感受、触觉信息处理等多种生理及行为背后的神经环路已通过活性依赖型报告基因结合光遗传学或感觉刺激等手段得到研究,但要全面明确这些环路的解剖结构与功能,需识别起始细胞的上下游细胞,而神经组织小区域内细胞类型的异质性,使得依赖麦胚凝集素(WGA)等蛋白质的经典示踪方法难以精确界定多数神经元环路。目前,基因修饰的狂犬病病毒可特异性感染表达禽肉瘤白血病病毒受体蛋白(TVA)的神经元,实现特定细胞类型的逆行跨单突触示踪,却缺乏能对基因定义细胞进行顺行示踪的类似工具,虽有腺相关病毒(AAV1)和基因修饰的黄热病毒(YFV)用于非特异性细胞的顺行示踪,但无法满足从基因定义细胞出发进行示踪的需求。
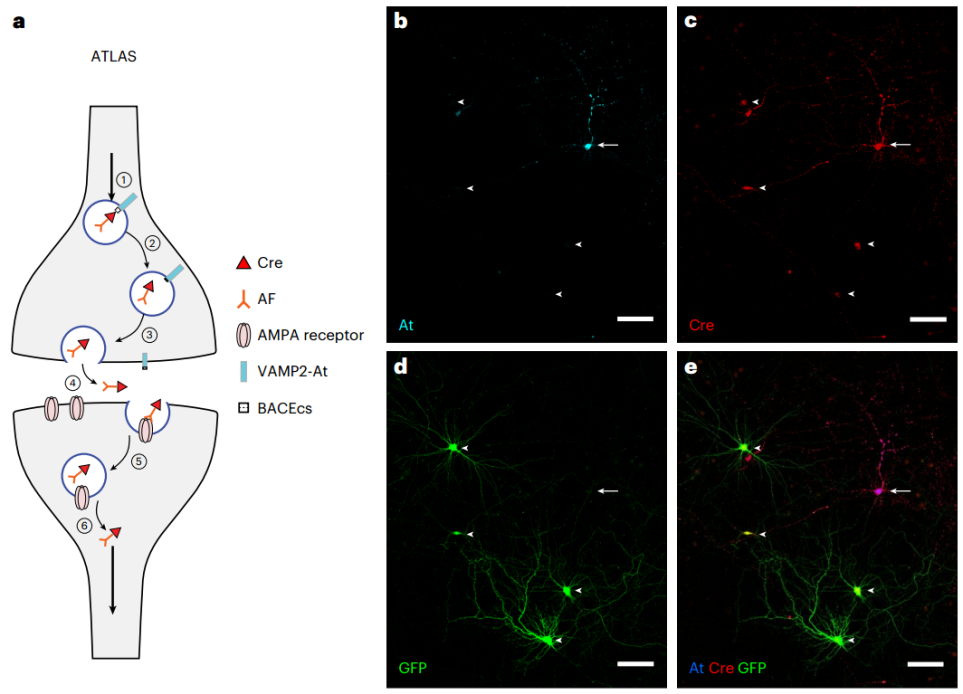
2025年5月,美国南加州大学Don B. Arnold团队在期刊Nature Methods上发表了题为“ATLAS: a rationally designed anterograde transsynaptic tracer”的研究论文,该研究开发了一种基于合理设计蛋白质ATLAS的顺行跨单突触示踪工具,可介导从基因定义细胞出发的顺行跨突触标记。ATLAS在神经元中表达时,会促使突触前膜释放包含两种功能成分的货物:一是抗体样蛋白AMPA.FingR(可特异性结合GluA1的N端——GluA1为AMPA型离子型谷氨酸受体的关键亚基);二是重组酶。该货物释放至突触间隙后,AMPA.FingR通过与突触后膜GluA1的N端结合形成复合物,进而介导整个货物被突触后细胞的内吞作用摄入;进入细胞后,重组酶在核定位信号引导下被输送至细胞核,最终通过切割报告基因中的特异性识别序列(如loxP位点),触发重组酶依赖的报告基因表达。在小鼠实验中,ATLAS能介导从随机或基因定义细胞出发的跨神经元单突触示踪,该示踪具有严格的顺行性、突触特异性、无毒性以及活动依赖性,可标记特定行为相关的活跃环路,同时其模块化组件可被独立替换或修饰,是一种可通过升级或改造来执行特定任务的多功能平台,为未来神经环路精细图谱的绘制及功能研究提供了强大且可扩展的有力工具。
枢密科技重磅推出神经环路示踪革新工具——ATLAS顺行跨单突触示踪系统!通过靶向模块升级彻底消除毒性,同时保持高效跨突触标记能力,严格限定顺行跨单突触示踪,无逆行跨突触干扰,具有活动依赖性特性,可广泛适用于多种类型的神经环路示踪。选择枢密科技,解锁神经环路示踪新维度!
实用Tips
1)病毒搭配方案:AAV-ATLASsnCre-WPRE-pA与AAV-DIO-mCherry/EGFP搭配使用,具体为上游脑区注射ATLASsnCre病毒,下游靶区注射DIO-mCherry/EGFP病毒。
2)效率优化策略:实验结果表明,当上游脑区混合注射AAV-ATLASsnCre-WPRE-PA与AAV-hSyn-HA-BACE-WPRE-PA(混合比例为BACE:ATLASsnCre=5:1),同时下游靶区注射DIO-mCherry/EGFP时,可显著提升跨单突触标记效率。
3)示踪层级限制:ATLAS系统仅能实现顺行跨单突触示踪,无法用于顺行跨多突触的研究。
4)方向特异性:ATLAS系统几乎无逆行跨突触标记,其跨单突触示踪具有严格的顺行性。
5)适用细胞类型:适用于兴奋性神经元的顺行跨单突触示踪。
6)活动依赖性特征:当上游注射区域的细胞被激活时,下游靶区的DIO-mCherry/EGFP荧光强度会显著增加。
1、AF的产生——一种GluA1 N端结合物
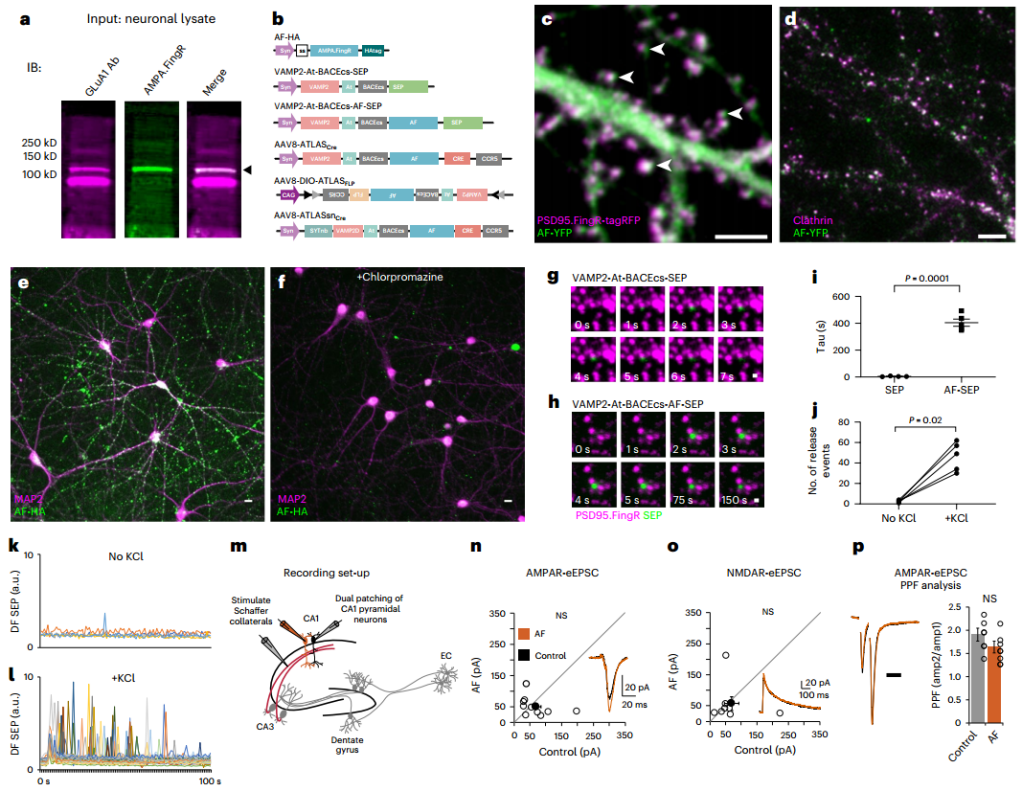
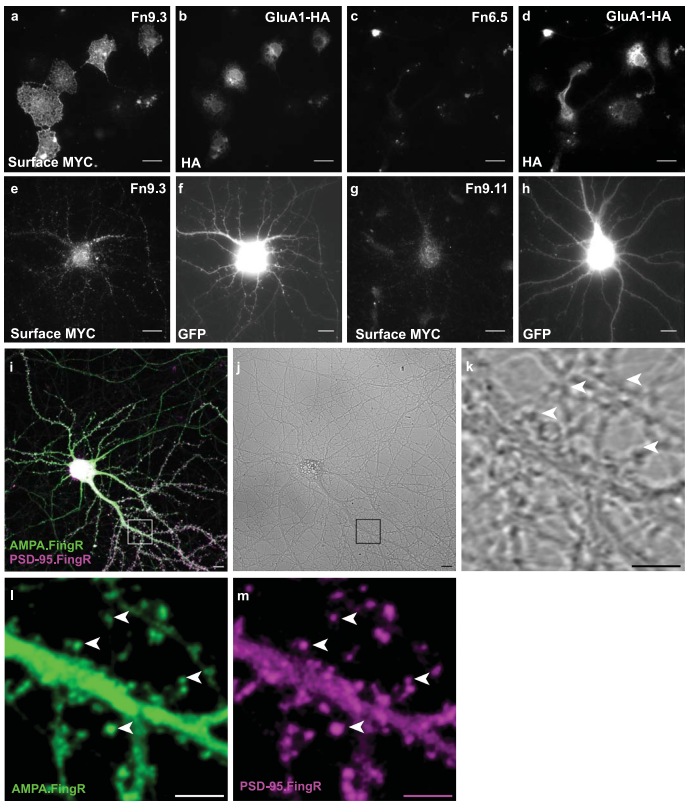
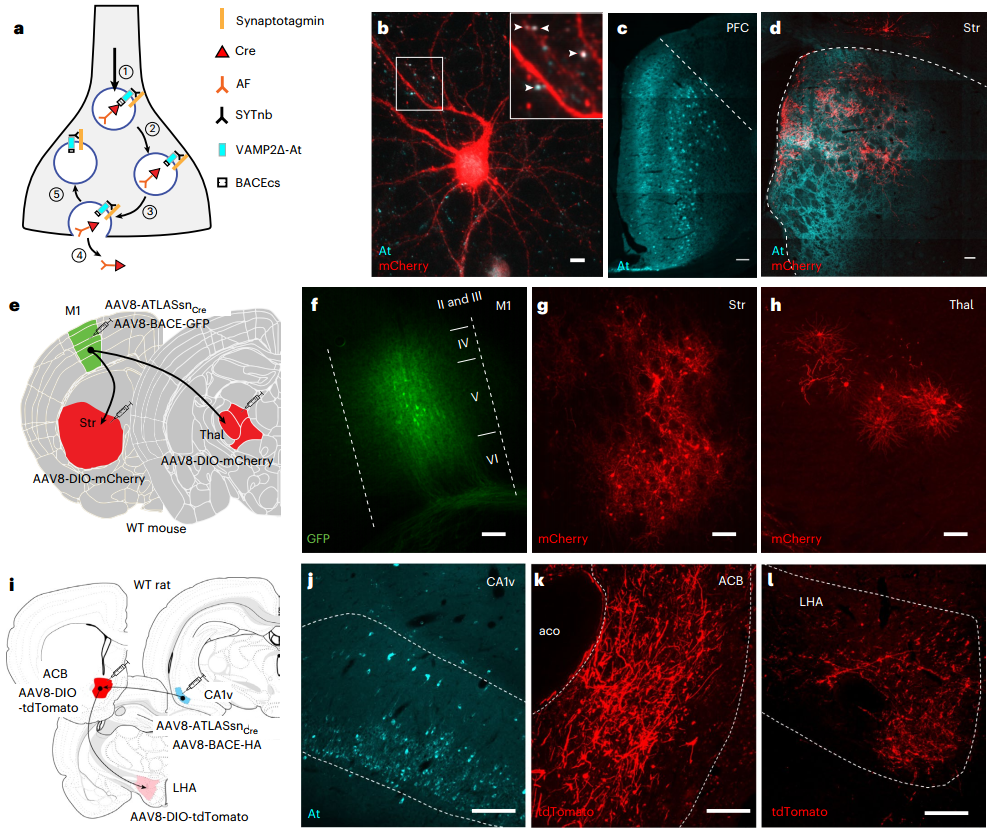
研究团队首先通过mRNA展示技术,从特定的mRNA-蛋白质融合文库中筛选出能与大鼠GluA1第1-394位氨基酸结合的结合物(纤维连接蛋白变体),在筛选获得的纤维连接蛋白变体中,Fn9.3克隆(命名为AMPA.FingR,简称AF)在原代皮层神经元中展现出最优的标记效果。为进一步表征AF,研究人员在原代皮质神经元中共表达其分泌型ss-AF-YFP(ss指GluA1信号序列)和突触后标记物PSD-95.FingR-tagRFP。结果显示AF与PSD-95.FingR-tagRFP部分重叠,提示其突触周围定位特征,同时,AF还与内源性网格蛋白共定位,提示其内吞区定位特征。进一步实验显示,外源性添加AF能使培养的皮质神经元被AF标记,而网格蛋白抑制剂氯丙嗪能显著抑制这一过程,证实AF的内化由内吞作用介导。
为实现跨突触示踪,研究人员构建了融合蛋白系统(ATLAS):将AF与突触前蛋白突触小泡蛋白2(VAMP2)的腔域融合,中间插入BACE1(β-分泌酶)切割位点(BACEcs)(形成VAMP2-BACEcs-AF),这样内源性BACE可将AF从VAMP2上切割下来,使其在胞吐时能从突触小泡腔中释放出来,此外还添加了ALFA标签(At)以标记VAMP2。为验证该策略,研究者用SEP(super ecliptic pHluorin,pH敏感荧光蛋白)替代AF构建了重组质粒(VAMP2-At-BACEcs-SEP),并与PSD-95.FingR-tagRFP在培养的皮质神经元中共表达。结果显示,在突触后位点对侧观察到符合突触小泡释放特征的短暂(约2-8秒)荧光爆发(SEP从酸性突触小泡释放到中性的突触间隙后,其荧光突然增强),表明VAMP2将SEP运送至突触小泡、BACE切割SEP并使其释放到突触间隙,证实了VAMP2靶向递送及BACE介导切割的可行性。
进一步用AF-SEP融合蛋白测试发现,释放后的荧光信号显著延长(405±60秒vs SEP的6±2秒),这与AF释放后结合到突触后膜并持续存在直至被内吞的特征相符。更重要的是,高钾去极化刺激使AF-SEP释放事件数量激增约15倍,表明AF-SEP是从突触小泡释放的。即AF-SEP从突触小泡释放后,穿过突触间隙,结合到相邻突触后神经元的突触周围区域,随后被内吞。此外,在海马CA1神经元的实验中,ss-AF-YFP的表达对AMPA受体、NMDA受体诱发的兴奋性突触后电流(eEPSCs)振幅及双脉冲易化均无显著影响,表明AF的表达不影响突触传递。
图1. ATLAS的释放与摄取机制
图2. AMPA.FingR的测试
2、ATLAS介导的顺行跨突触示踪:基因靶向性与效率优化
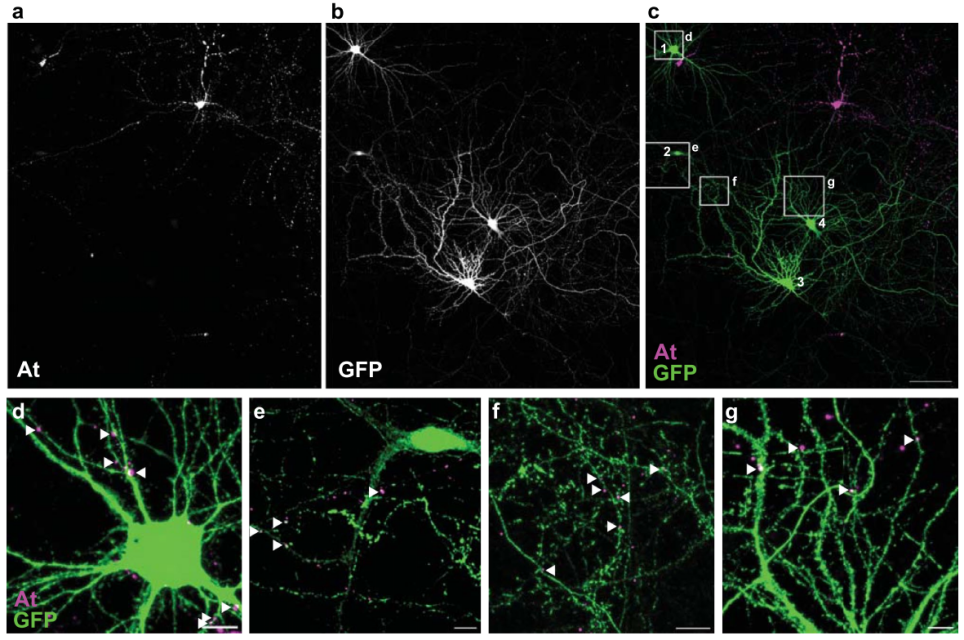
在成功构建AF及其递送机制的基础上,研究者进一步开发了完整的顺行跨突触示踪工具ATLAS。首先,将报告蛋白SEP替换为Cre重组酶,构建了ATLASCre(VAMP2-At-BACEcs-AF-Cre),并在ATLAS蛋白中引入含强核定位信号的CCR5 DNA结合域,以确保AF-Cre能被递送至突触后细胞核,从而实现对突触后细胞的遗传学调控。为验证该策略,研究人员将ATLASCre转染至已感染AAV8-DIO-GFP的原代培养细胞中,成功观察到未被At标记却表达GFP和Cre的神经元,且At标记(表明是起始细胞)的突触前结构与GFP+神经元的树突紧密相邻或重叠,表明ATLASCre可介导跨突触示踪及Cre的核转位,并为突触后细胞提供遗传学调控的可能。
图3. ATLAS跨突触转运及体外测试示意图
图4. ATLAS在培养体系中的跨突触标记
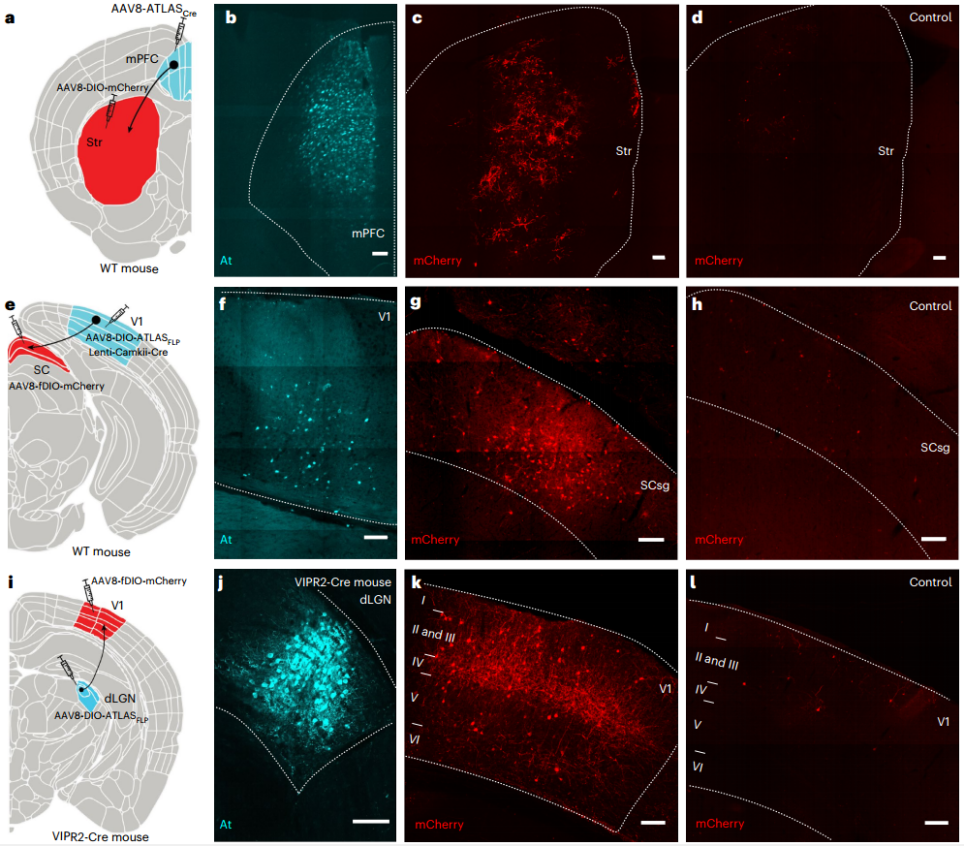
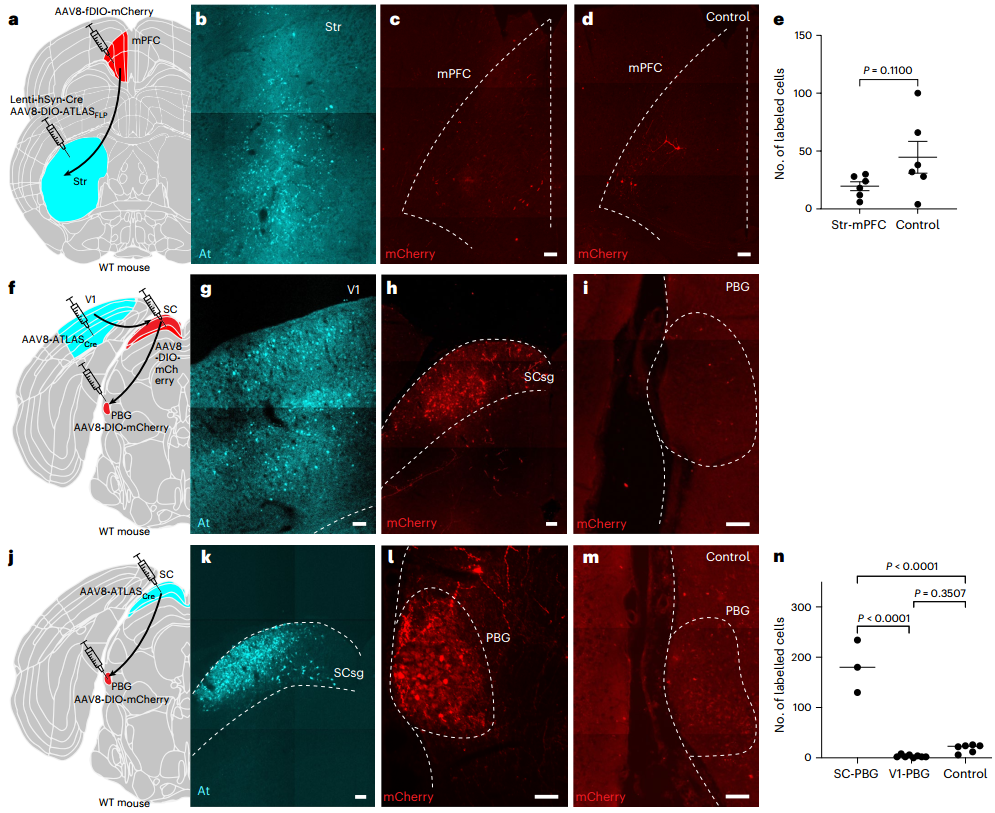
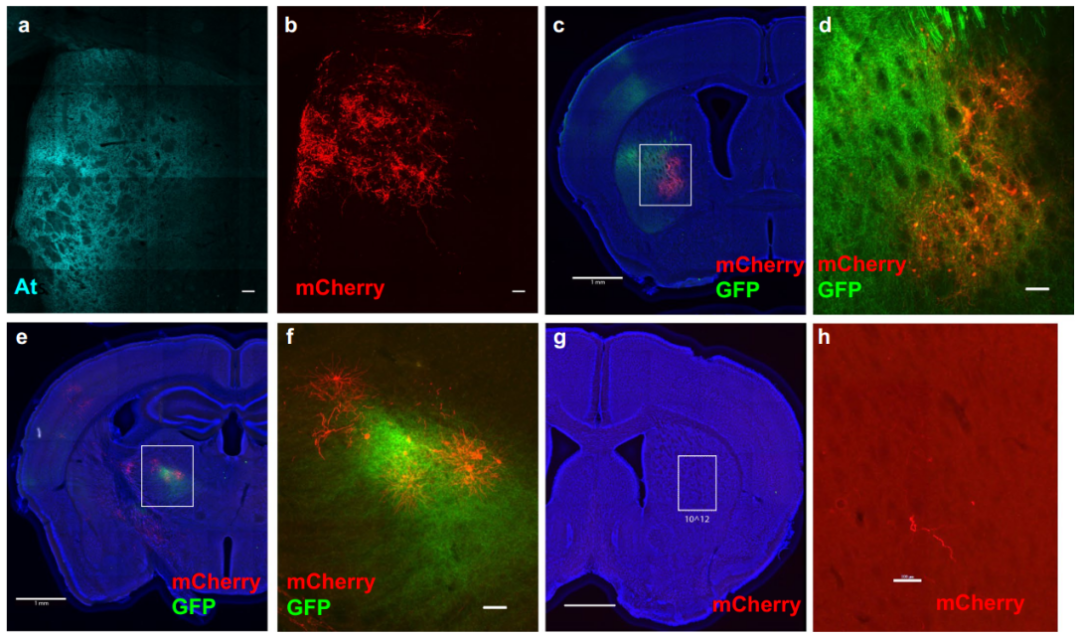
为验证其体内效能,研究者将AAV8-ATLASCre注射至小鼠内侧前额叶皮层(mPFC),同时将AAV8-DIO-mCherry注射至其下游脑区纹状体(Str)。一周后,在mPFC中检测到了At染色,并且在Str观察到大量mCherry+细胞,而对照组(无ATLASCre)则无此现象,证实ATLAS在体内可实现顺行跨突触标记。
为实现基因定义示踪,研究者构建了Cre依赖性表达的AAV8-DIO-ATLASFLP(携带FLP重组酶)。在野生型小鼠中,将表达Cre的慢病毒(Lenti-CaMKII-Cre,特异性靶向兴奋性神经元)与AAV8-DIO-ATLASFLP共注射至初级视皮层(V1),并将FLP依赖的AAV8-fDIO-mCherry注射至其严格顺行投射的下游上丘(SC)。结果除在V1中观察到At染色外,在SC检测到大量mCherry+细胞,显著高于对照组(无ATLASFLP)。该策略的优势在于慢病毒无内在的跨神经元顺行或逆行转运能力,不会干扰结果的解读。研究人员进一步在VIPR2-Cre转基因鼠(背外侧膝状核(dLGN)神经元中表达Cre)的dLGN注射AAV8-DIO-ATLASFLP,并在其投射脑区V1注射AAV8-fDIO-mCherry,结果在dLGN中特异性检测到At染色,并成功在V1的IV层(主要投射层)及II/III和V层观察到符合解剖学特征的mCherry标记。这些结果共同证明ATLAS在野生型和转基因小鼠中均能有效介导从基因定义神经元出发的顺行跨突触示踪。
图5. ATLAS在体内介导的顺行跨突触示踪
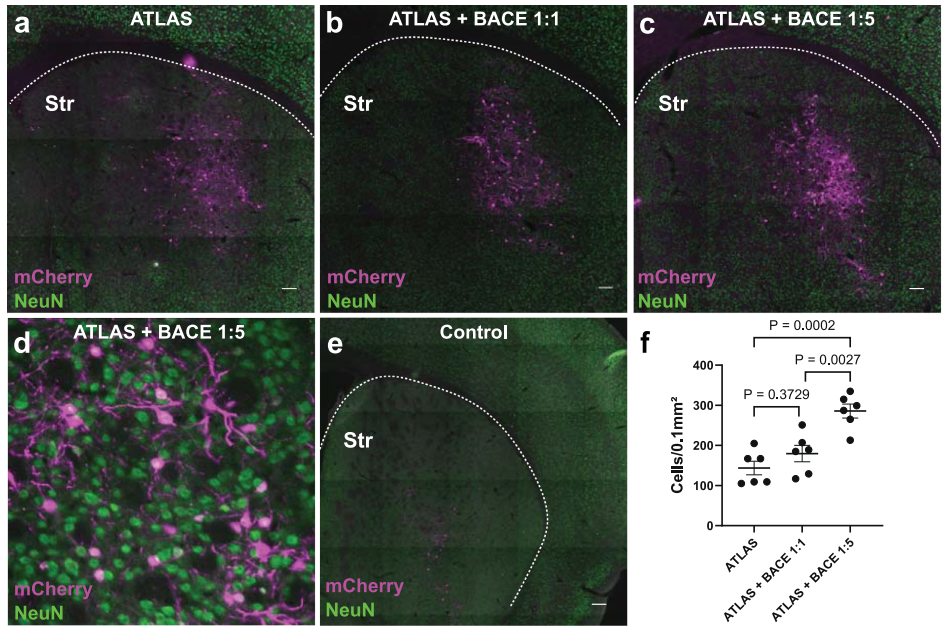
最后,针对初始标记效率(标记mPFC→Str通路约144±17个mCherry+细胞),研究者探索了优化策略。推测内源性BACE酶的量可能不足以将所有AF-Cre从VAMP2上切割下来,故尝试共表达外源BACE。当以1:1比例共注射AAV8-BACE-HA与AAV8-ATLASCre至mPFC时,Str中mCherry标记细胞数未显著增加(180±20个)。然而,当以5:1(病毒颗粒数)的比例注射AAV8-BACE-HA和AAV8-ATLASCre时,Str的mCherry+细胞数显著增加至286±17个,效率提升近一倍,达到约10%的突触后神经元标记率,与已报道的YFV顺行示踪工具效率相当。综上,通过补充外源BACE可有效提升ATLAS的跨突触标记能力。
图6. 通过共表达BACE提高ATLAS的效率
3、ATLAS示踪的跨突触特性、无神经毒性及电生理学特异性验证
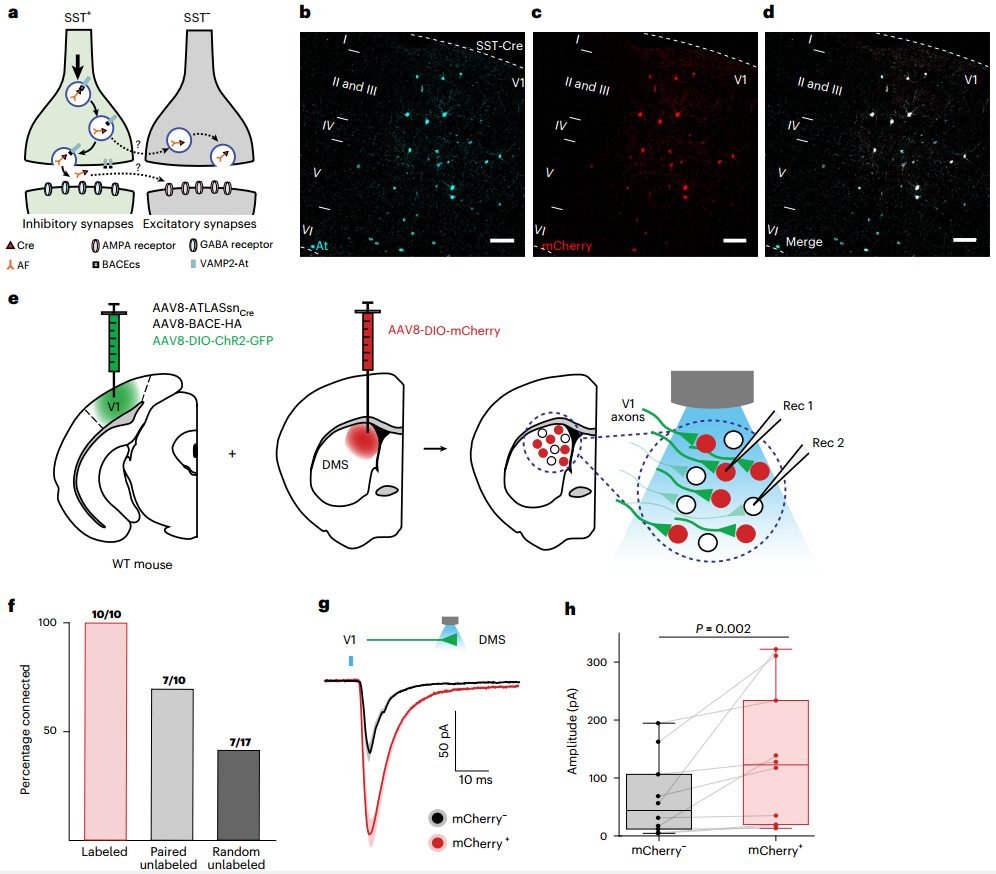
为明确ATLAS跨突触标记的突触特异性,研究团队利用SST-Cre小鼠(生长抑素(SST)阳性抑制性中间神经元特异性表达Cre酶)进行实验:在SST-Cre小鼠的V1区注射AAV8-DIO-ATLASFLP与AAV8-fDIO-mCherry。由于抑制性突触缺乏GluA1受体(ATLAS内吞的必需靶点),理论上出现任何跨神经元标记都必然是非突触性的。结果显示,两周后V1中几乎所有表达mCherry的神经元都被At共标记,证明ATLAS仅通过蛋白质的跨突触转运对突触后神经元进行标记。
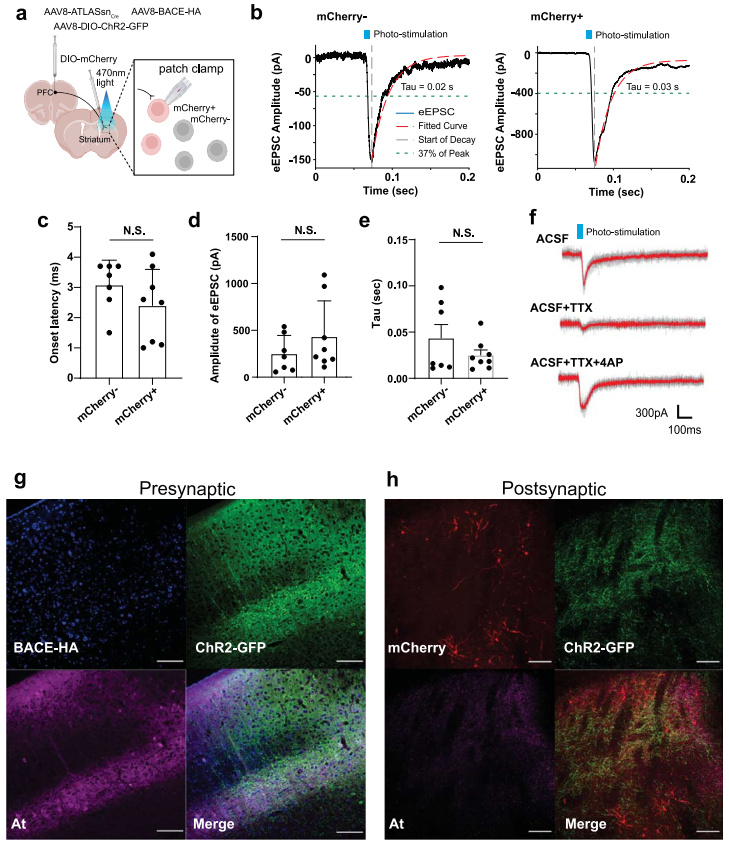
为评估ATLAS对突触传递的影响,研究人员在mPFC共注射AAV8-ATLASsnCre(优化版ATLAS)、AAV8-BACE-HA及AAV8-DIO-ChR2-GFP,并向Str注射AAV8-DIO-mCherry。通过测量Str细胞中470 nm光诱发的mCherry+(ATLAS系统标记)与mCherry-(未被ATLAS系统标记)细胞的eEPSC,发现两组光诱发eEPSC的幅度、潜伏期及时间常数均无显著差异。此外,团队结合河豚毒素(TTX,阻断动作电位和多突触传递)与4-氨基吡啶(4-AP,可阻断K+通道并促进突触前末梢直接激活)验证了电流为单突触性质。以上结果表明ATLAS不影响突触传递功能且无毒性。
为进一步验证示踪特异性,研究者在V1共注射了AAV8-ATLASsnCre和AAV8-BACE-HA,并在背内侧纹状体(DMS)注射了AAV8-DIO-mCherry/tdTomato。两周后在V1与首次注射相同的区域注射了光遗传学病毒AAV8-DIO-ChR2-GFP,以调控ATLAS标记的突触前输入,同时最大限度地减少ATLAS与ChR2之间的表达竞争。在病毒注射4周后制备的急性脑切片中,对同一视野(100×100 μm)内ATLAS标记(mCherry+)与未标记(mCherry-)DMS神经元(共10对)进行光刺激记录。结果显示,ATLAS标记的突触后细胞不仅连接概率(10个标记细胞均记录到电流)显著高于未标记细胞(10个未标记细胞中仅有7个记录到电流),其光诱发EPSC的振幅和总电荷也显著大于未标记细胞,且在10对细胞中,所有标记细胞记录到的电流振幅均大于未标记细胞。以上结果表明ATLAS系统能够标记与起始神经元有强突触连接的下游细胞,且连接越强,标记越容易。
图7. ATLAS不介导非突触的跨神经元标记,且跨突触标记具有特异性
图8. ATLAS示踪不影响突触传递功能
4、ATLAS具备严格顺行性、跨单突触特性及活动依赖性
为验证ATLAS无逆行标记特性,研究人员将AAV8-DIO-ATLASFLP与不会逆行传播的慢病毒Lenti-hSyn-Cre注射到Str,并在其上游脑区mPFC注射AAV8-fDIO-mCherry(mPFC到Str的投射是单向的)。结果显示,尽管Str区中有大量At标记(表明ATLAS高水平表达),但在mPFC中未检测到高于背景水平的标记,证明ATLAS不会介导逆行跨突触传递。
为验证其跨单突触特性,团队针对V1→SC→二叠体旁核(PBG)这一多突触通路展开实验:将AAV8-ATLASCre注入V1,向SC及PBG注射AAV8-DIO-mCherry。结果仅在SC检测到大量mCherry+细胞,而PBG中没有显著高于背景的标记。反向对照中,将AAV8-ATLASCre直接注入SC后,PBG出现显著的mCherry标记,并且显著高于背景和从V1到PBG的标记。证实ATLAS介导的跨突触标记是严格顺行且单突触的。
图9. ATLAS的跨突触标记具有严格的顺行性和单突触性
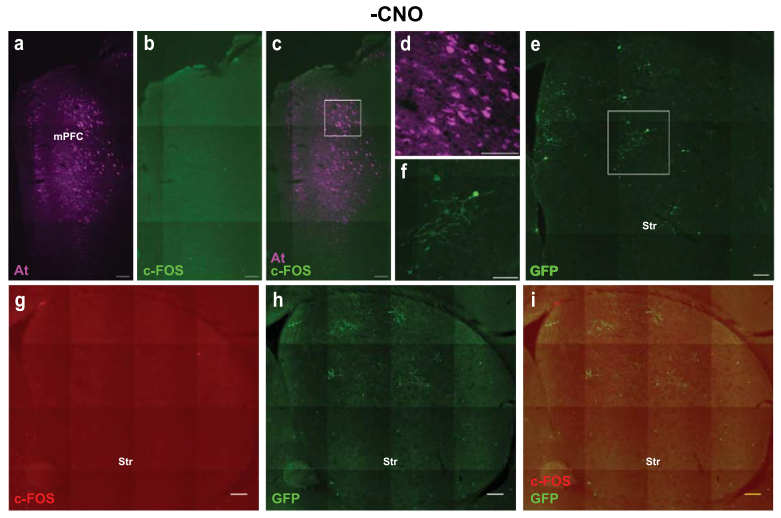
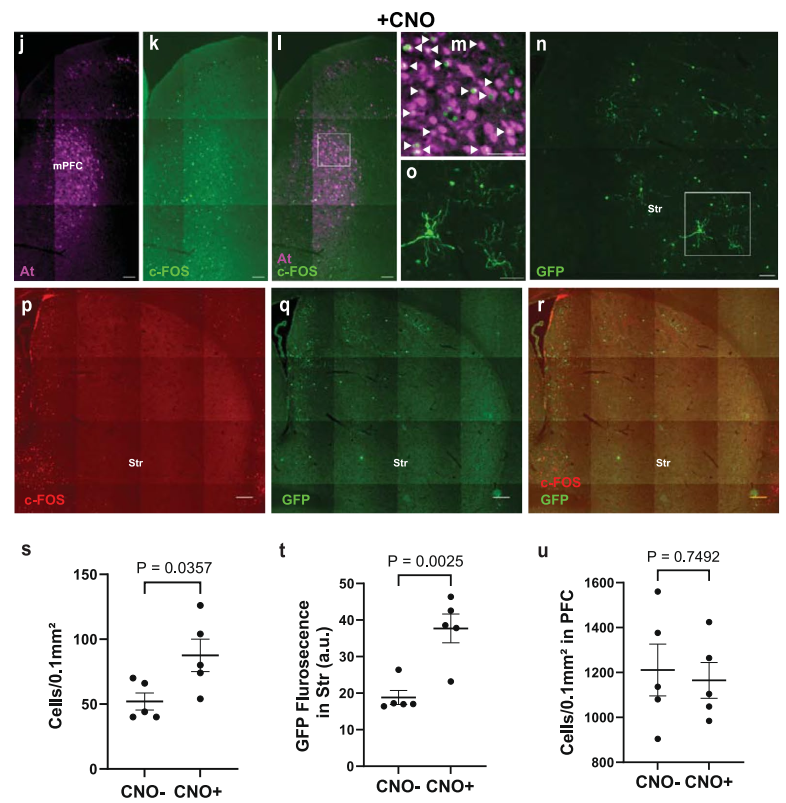
为检验ATLAS在体内跨突触标记的活动依赖性,团队在mPFC共注射AAV8-ATLASsnCre与化学遗传激活病毒AAV8-hHM3Dq-mCherry,并在Str注射AAV8-DIO-GFP。结果显示,连续腹腔注射CNO的小鼠mPFC区存在大量c-Fos标记且与At共定位,未注射CNO的小鼠mPFC中仅检测到极少的c-Fos标记;且注射CNO的小鼠Str的GFP+细胞数及总荧光强度均显著高于未激活组(未注射CNO)。证实ATLAS的跨突触标记效率依赖于起始细胞的活动状态。
图10. ATLAS的体内跨突触标记具有活动依赖性
5、ATLAS的组件优化:消除毒性、环路通用性及跨物种适用性
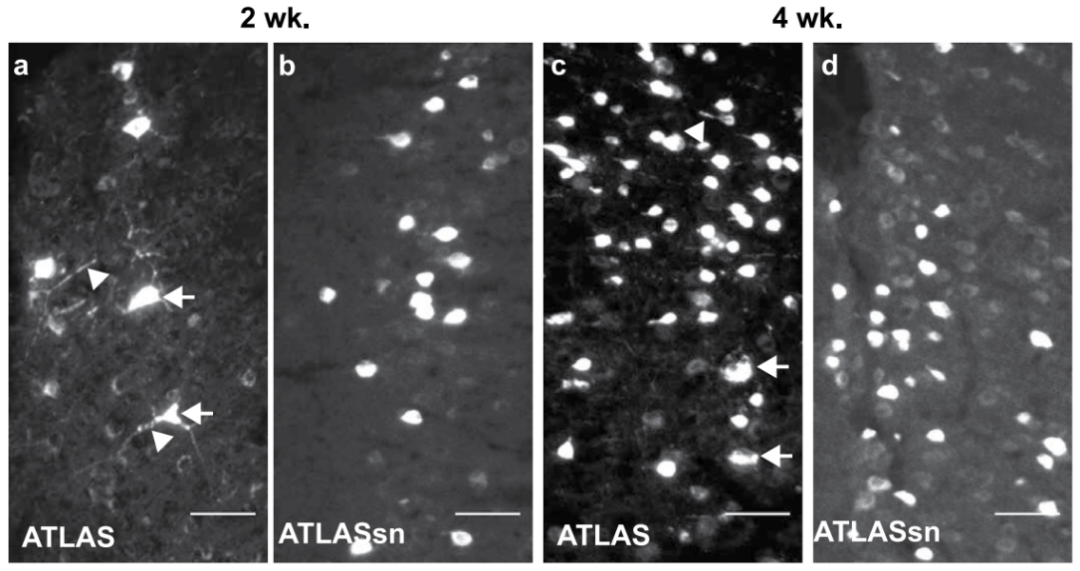
研究团队利用ATLAS的模块化特性进一步对其进行了优化,针对VAMP2高表达产生的毒性(在mPFC表达第2和4周时),研究者将VAMP2胞质结构域替换为靶向突触结合蛋白1的纳米抗体(SYTnb),构建出改良版ATLASsnCre(SYTnb-VAMP2Δ-At-BACEcs-AF-Cre)。该版本在小鼠mPFC中表达2或4周时,均未观察到毒性。体外实验中,ATLASsnCre成功介导突触后mCherry标记,且At信号定位于突触前末梢。体内实验进一步证实:在mPFC→Str通路中,ATLASsnCre联合外源BACE可实现高效跨突触标记,且Str区mCherry+细胞与突触前末梢At标记几乎完全共定位;同时在初级运动皮层(M1)→Str/丘脑(Thal)环路中,Str与Thal均检测到GFP(BACE标记)与mCherry共标细胞,符合跨突触标记特征。以上结果表明,改良版ATLASsnCre不仅消除了毒性,且跨突触标记特征未受影响。
图11. ATLASsn的跨突触示踪
图12. ATLAS具有毒性,而ATLASsn无毒性
图13. ATLASsnCre从皮层到Str及Thal的跨突触标记
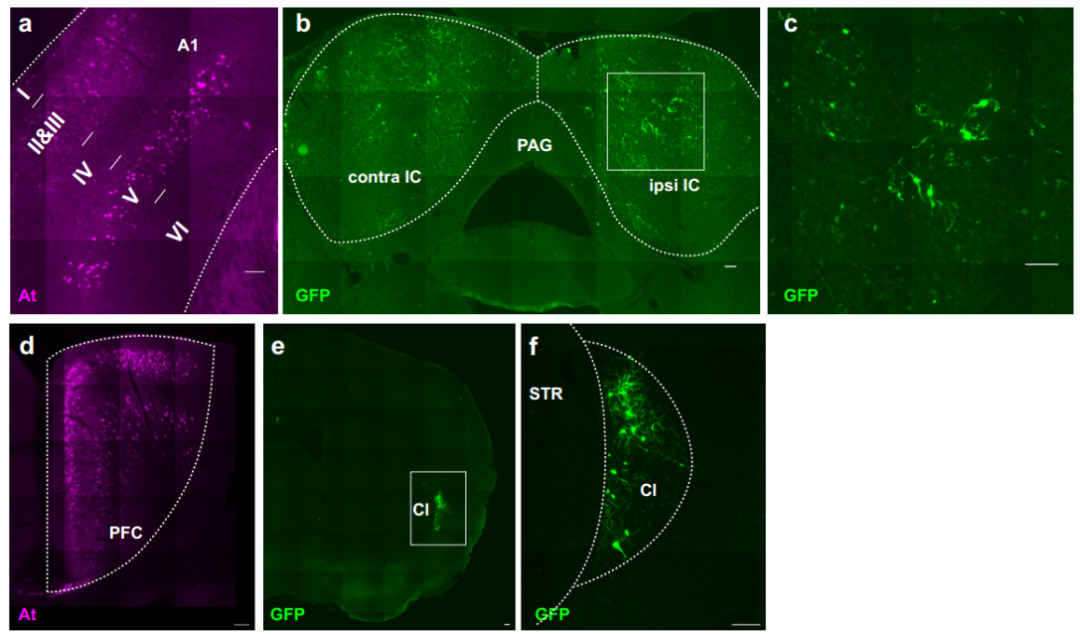
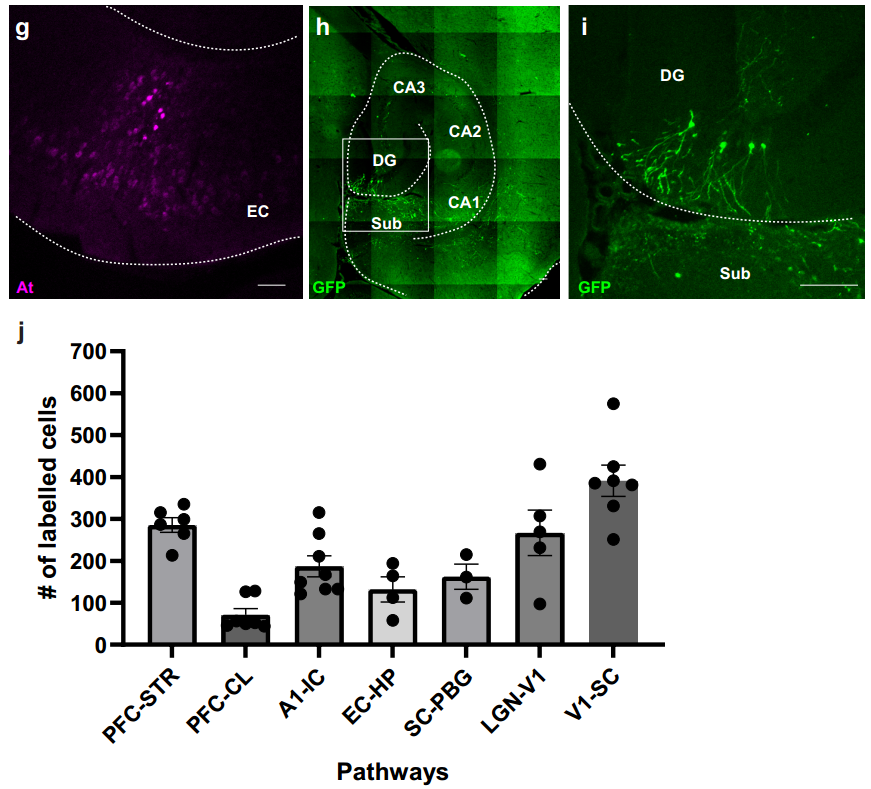
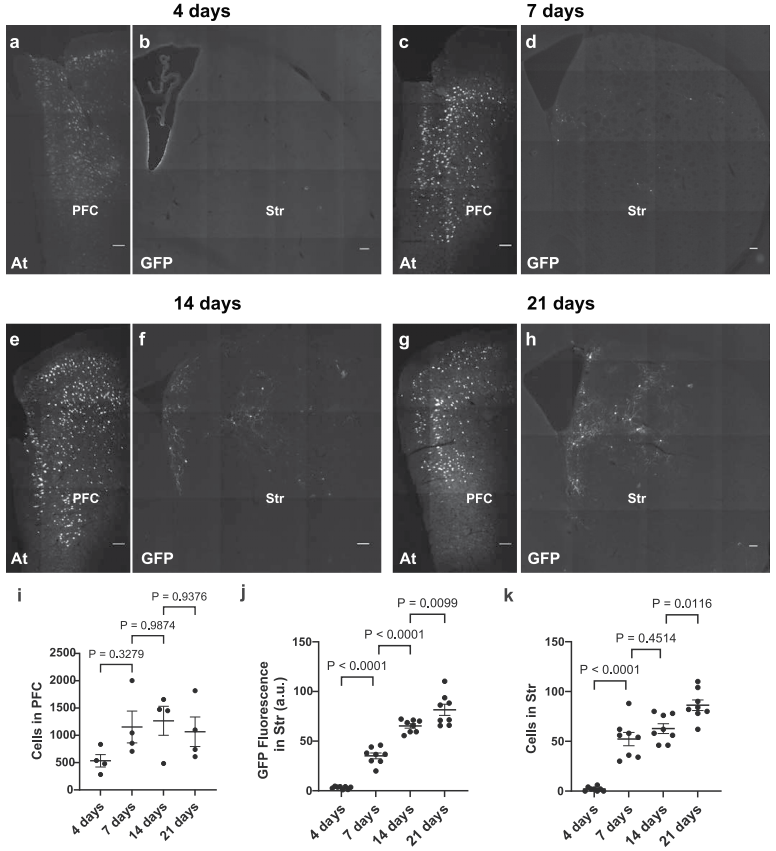
为探究ATLASsn的通用性,团队利用ATLASsn成功标记三条新环路:听觉皮层(A1)→下丘、mPFC→屏状核、内嗅皮层→海马,表明ATLASsn可有效追踪多种不同神经环路。此外,为明确跨突触标记的时间进程,团队在小鼠mPFC共表达AAV8-ATLASsnCre与AAV8-BACE-HA,在Str表达AAV8-DIO-GFP,分别于4、7、14和21天后灌注小鼠并对突触前及突触后标记进行染色。结果显示,ATLAS突触后标记在注射后4-7天开始出现,之后细胞数量的增加速率减慢,但总荧光强度呈相对线性增长;相比之下,突触前标记细胞数量在第1周后保持稳定。
图14. ATLASsnCre从皮层到皮层下区域的跨突触标记
图15. mPFC至Str跨突触示踪的时间进程
最后,研究还将ATLAS应用拓展至大鼠模型。团队向大鼠腹侧海马CA1区注射AAV8-ATLASsnCre和AAV8-BACE-HA,并在其投射的伏隔核(ACB)注射AAV8-DIO-tdTomato,结果在CA1区观察到At染色,ACB中出现大量tdTomato标记细胞;同时,外侧下丘脑区(LHA,接收ACB轴突投射)的轴突也出现tdTomato标记。证实ATLAS在大鼠中同样能有效介导跨突触标记,展现出良好的跨物种适用性。(见图11)
总结
本研究开发的ATLAS作为一种合理设计的蛋白质工具,通过明确的分子机制实现了从基因定义神经元出发的严格顺行、单突触、活动依赖性跨突触标记,突破了现有病毒工具(如狂犬病毒、YFV、AAV1等)在作用机制不明确、存在逆行标记、多突触扩散及神经毒性等方面的局限。
然而ATLAS当前应用仍存在一项技术约束:需预先确定突触后靶区并表达重组酶依赖的报告基因。展望未来,ATLAS的模块化设计赋予其强大的可拓展性:通过替换AF靶向模块(如适配GABA、乙酰胆碱乃至多巴胺/血清素等神经调质受体),有望实现对各类神经递质环路的普适性标记;结合其活动依赖性特征,有望实现特定刺激响应环路的标记,甚至追踪记忆形成、突触修剪等生理过程中的环路动态变化。作为一种基于蛋白质的遗传靶向顺行示踪平台,ATLAS为解析神经环路的解剖连接与功能编码提供了不可替代的新范式。
我司可提供ATLAS顺行跨单级突触病毒工具,列表如下:


详情欢迎扫码咨询
微信号:BrainVTA2020

市场:027-65023363 行政/人事:027-62439686 邮箱:marketing@brainvta.com 客服:18140661572(活动咨询、售后反馈等)
销售总监:张经理 18995532642 华东区:陈经理 18013970337 华南区:王经理 13100653525 华中/西区:杨经理 18186518905 华北区:张经理 18893721749
地址:中国武汉东湖高新区光谷七路128号中科开物产业园1号楼
Copyright © 武汉枢密脑科学技术有限公司. All RIGHTS RESERVED.
鄂ICP备2021009124号 DIGITAL BY VTHINK