2025-07-01 阅读量:1833
克罗恩病(CD)是一种慢性胃肠道炎症疾病,病因复杂。学界公认,遗传因素、环境因素与微生物群之间的相互作用会引发过度活跃的黏膜免疫反应和胃肠上皮屏障损伤。在生理条件下,上皮层调控着多种功能,包括营养循环、黏液分泌、宿主-微生物互作以及黏膜免疫应答。然而,过度激活的免疫细胞和上皮细胞会通过促炎细胞因子的诱导,产生过量活性氧(ROS)。持续的ROS过载将导致肠道屏障功能受损、DNA损伤及细胞死亡失调。肠上皮细胞中失调的坏死性细胞死亡会进一步加剧促炎性免疫反应,并对上皮组织造成更严重的损害。
斯钙素-1(STC1)是一种糖蛋白,影响多种功能,包括血管生成、肿瘤发生和女性生殖。STC1在炎症期间显著上调,且具有应激响应性,在过氧化氢(H2O2)刺激下会增加。据报道,STC1可通过增加线粒体解偶联蛋白2(UCP2)和抑制ROS诱导的细胞凋亡来减轻氧化应激。然而,STC1在体外和体内均能通过增加ROS和诱导细胞死亡介导心脏毒性。因此,现有文献表明STC1可能作为应激响应因子和凋亡介质发挥作用。氧化应激可诱导多种类型的细胞死亡,如铁死亡、铜死亡和PARP1依赖性细胞死亡[parthanatos:PARP1(聚(腺苷二磷酸/ADP-核糖)聚合酶-1)依赖性的程序性坏死性细胞死亡]。然而,STC1在这些新发现的细胞死亡类型中的功能尚需进一步研究。
在多种形式的细胞死亡中,关注PARP1依赖性细胞死亡的研究较少。这种形式的细胞死亡,由损伤刺激(如氧化应激和炎症刺激)下PARP1的过度激活所启动。过度激活的PARP1会产生PAR[聚(ADP-核糖)]聚合物,这些聚合物从细胞核释放到细胞质中。PAR与凋亡诱导因子(AIF)结合并将其从线粒体中释放出来。随后,AIF与巨噬细胞迁移抑制因子(MIF)(一种DNA核酸酶)结合,并易位至细胞核,导致基因组DNA切割(断裂)。作为一种程序性坏死性细胞死亡,PARP1依赖性细胞死亡的发生不依赖于促凋亡的caspase级联反应,并会导致DNA断裂和细胞膜完整性丧失。因此,PARP1依赖性细胞死亡在多种疾病中表现出促炎特征,包括银屑病和脂肪性肝炎。此外,Parp1基因敲除小鼠表现出核因子-κB(NF-κB)活化缺陷和对炎症的抵抗性,包括对脂多糖(LPS)诱导的脓毒性休克和过氧亚硝酸盐诱导的关节炎的抵抗。更重要的是,抑制Parp1能显著减轻葡聚糖硫酸钠(DSS)诱导的结肠炎小鼠模型及白细胞介素-10(IL-10)缺陷的自发性结肠炎小鼠模型中的结肠炎症程度,并保护肠道屏障。基于现有证据,STC1和PARP1在CD发病机制中的作用仍不清楚。
近期,中山大学附属第一医院张盛洪主任医师团队在Advanced Science发表题为“Stanniocalcin-1 Promotes PARP1-Dependent Cell Death via JNK Activation in Colitis”研究论文。本研究中,作者探究了PARP1依赖性细胞死亡是否在CD氧化损伤的发病机制中起作用。此外,还阐明了STC1和PARP1介导CD中氧化应激诱导的结肠上皮损伤和PARP1依赖性细胞死亡的潜在机制。结果显示,在CD患者及化学诱导小鼠结肠炎模型的结肠炎症黏膜中,STC1表达显著增加。对PARP1依赖性细胞死亡严重程度和促炎细胞因子表达的评估表明,肠道特异性Stc1基因敲除(Stc1INT-KO)小鼠对DSS诱导的结肠炎具有抵抗力,并表现出较低的疾病严重程度。过表达STC1的细胞表现出更高程度的PARP1依赖性细胞死亡和促炎细胞因子表达,而STC1基因敲除的细胞则表现出较低程度的PARP1依赖性细胞死亡。免疫共沉淀(co-IP)、质谱和蛋白组学分析表明,STC1与PARP1相互作用,并通过PARP1-JNK相互作用激活JNK信号通路。此外,抑制PARP1和JNK可减轻由STC1过表达引发的PARP1依赖性细胞死亡和炎症损伤。最后,通过腺相关病毒(AAV)恢复Stc1和Parp1的表达,结果显示,Stc1和Parp1的过表达会加重Stc1INT-KO小鼠中DSS诱导的结肠炎。总的来说,在CD发病机制中,STC1通过STC1-PARP1-JNK相互作用及其后续的JNK通路激活,介导了氧化应激相关的PARP1依赖性细胞死亡并加剧了炎症反应。

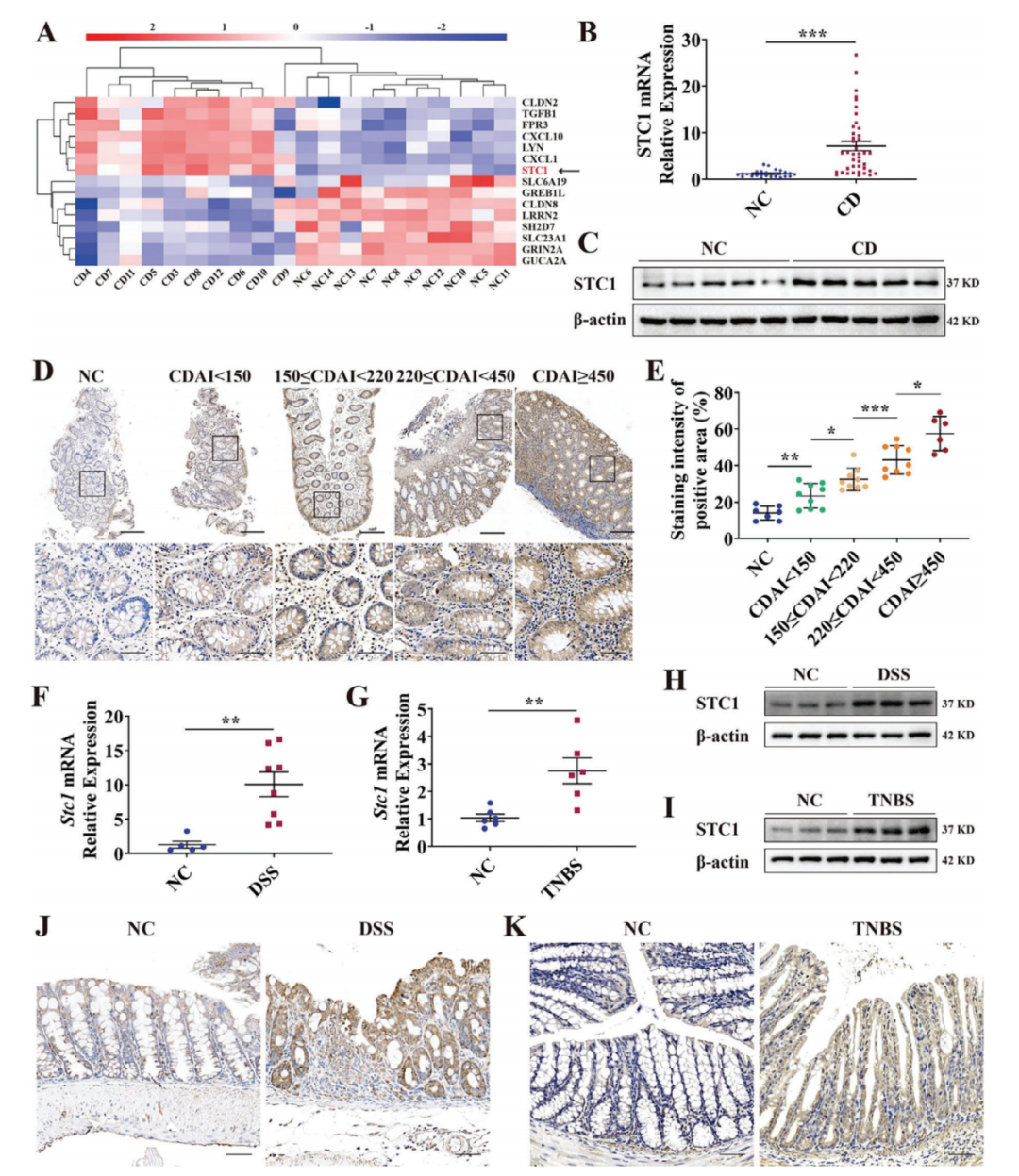
1.STC1在CD患者和化学诱导的小鼠结肠炎模型结肠黏膜中表达上调
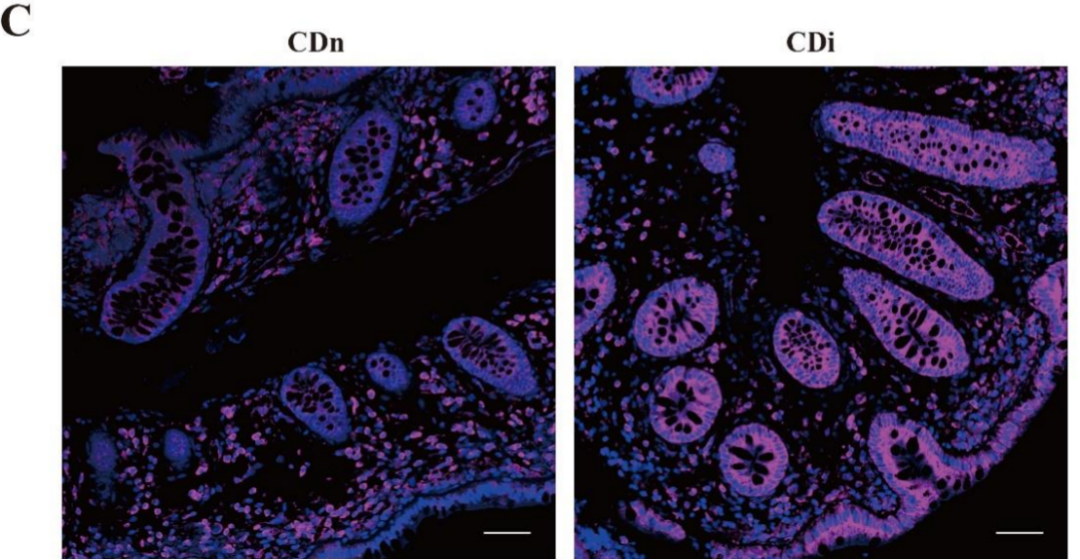
为鉴定CD中的分子因子,作者对10名CD患者的结肠炎症黏膜和10名健康对照者的结肠正常黏膜进行了转录组测序(GSE230113),结果显示,CD患者的结肠黏膜中检测到更高水平的STC1 mRNA表达。这些结果在扩大样本量(CD组,n=41;NC组,n=25)后得到证实:与未发生炎症反应的健康黏膜相比,STC1 mRNA在CD患者结肠炎症黏膜中显著上调。此外,STC1蛋白的丰度在CD患者的结肠组织中亦显著增加。免疫组化(IHC)显示STC1的表达水平随着CD活动指数(CDAI)的增加而升高。免疫荧光染色显示,STC1在上皮细胞和免疫细胞中均有表达。然而,其在CD患者炎症和相对正常结肠组织的免疫细胞中的表达相似。相比之下,与CD患者相对正常的结肠组织相比,STC1在炎症结肠组织的上皮细胞中表达上调。因此,研究人员在后续分析中重点关注了STC1在上皮细胞中的功能。
研究者还在DSS和TNBS(2,4,6-三硝基.苯磺酸)诱导的急性结肠炎小鼠模型[造摸方法:1.喂食2.5%(w/v)DSS溶液一周;2.结肠内注射2.5%(w/v)TNBS溶液]中检测了Stc1的表达。结果显示,Stc1 mRNA在DSS和TNBS诱导的小鼠结肠炎模型中均上调。类似地,两种结肠炎模型的结肠组织中STC1蛋白水平均升高。
图1.STC1在CD患者和化学诱导的结肠炎小鼠模型结肠黏膜中表达增加
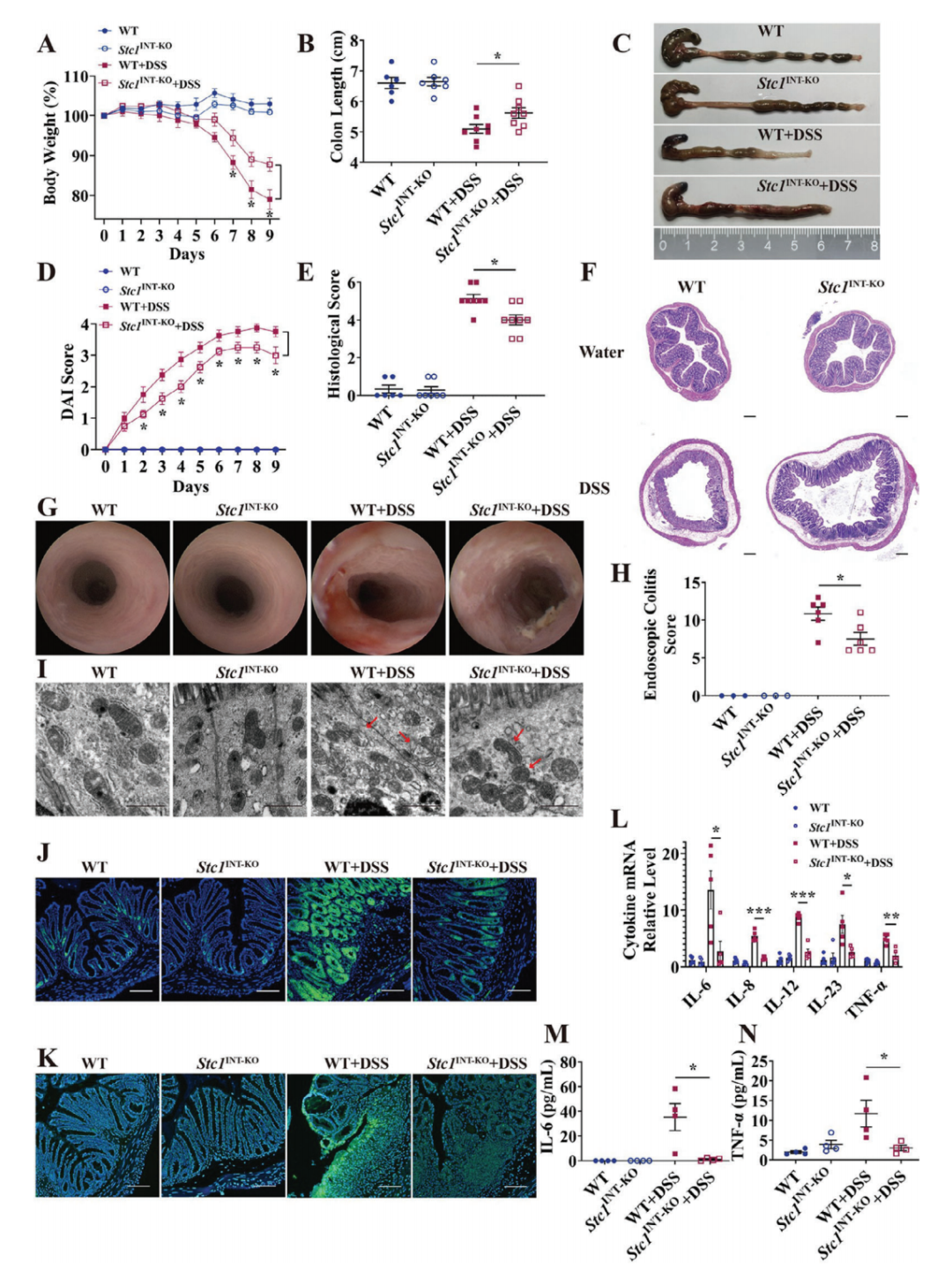
2.Stc1缺乏减轻DSS诱导的小鼠急性结肠炎
在DSS和TNBS诱导的小鼠结肠炎模型中,产生了大量的活性氧(ROS)。本研究使用DSS在小鼠结肠中诱导氧化损伤和炎症损伤。为研究Stc1在体内的功能,研究人员构建了肠道特异性Stc1基因敲除小鼠(Stc1flox/flox:Villin Cre+,Stc1INT-KO)及其同窝对照小鼠(Stc1flox/flox,WT)。结果显示,与WT+DSS小鼠相比,Stc1INT-KO+DSS小鼠表现出更少的体重减轻、更长的结肠长度、更低的疾病活动指数(DAI)评分、更轻的结肠上皮组织学损伤以及更低组织学评分。小鼠内镜检查显示,Stc1INT-KO+DSS小鼠的结肠壁仅观察到轻度损伤;而WT+DSS小鼠则表现出明显的血管模式改变、黏膜颗粒感增加和肠壁增厚,这些变化通过内镜结肠炎评分进行了量化。透射电子显微镜(TEM)进一步显示,WT+DSS小鼠的结肠上皮细胞中线粒体变形肿胀、嵴断裂;而Stc1INT-KO+DSS小鼠结肠上皮细胞中的线粒体损伤较轻。此外,研究人员采用组蛋白H2A.X变体磷酸化(γ-H2AX)来显示DNA双链断裂(DSB)。结果显示,Stc1INT-KO+DSS小鼠结肠上皮中产生的γ-H2AX水平较低,表明其DNA DSB频率更低。同时,使用PAR[聚(ADP-核糖)]生成来评估PARP1依赖性细胞死亡的程度,其在Stc1INT-KO+DSS小鼠中显著下调,表明Stc1缺乏改善了DSS诱导的小鼠结肠炎中的PARP1依赖性细胞死亡。促炎细胞因子,包括IL-6、IL-8、IL-12、IL-23和肿瘤坏死因子(TNF)-α,在WT+DSS小鼠的结肠组织中显著上调,但这种效应在Stc1INT-KO+DSS小鼠中被削弱。血清IL-6和TNF-α水平在WT+DSS和Stc1INT-KO+DSS小鼠中也表现出相似的模式。
图2.Stc1缺失减轻DSS诱导的小鼠急性结肠炎
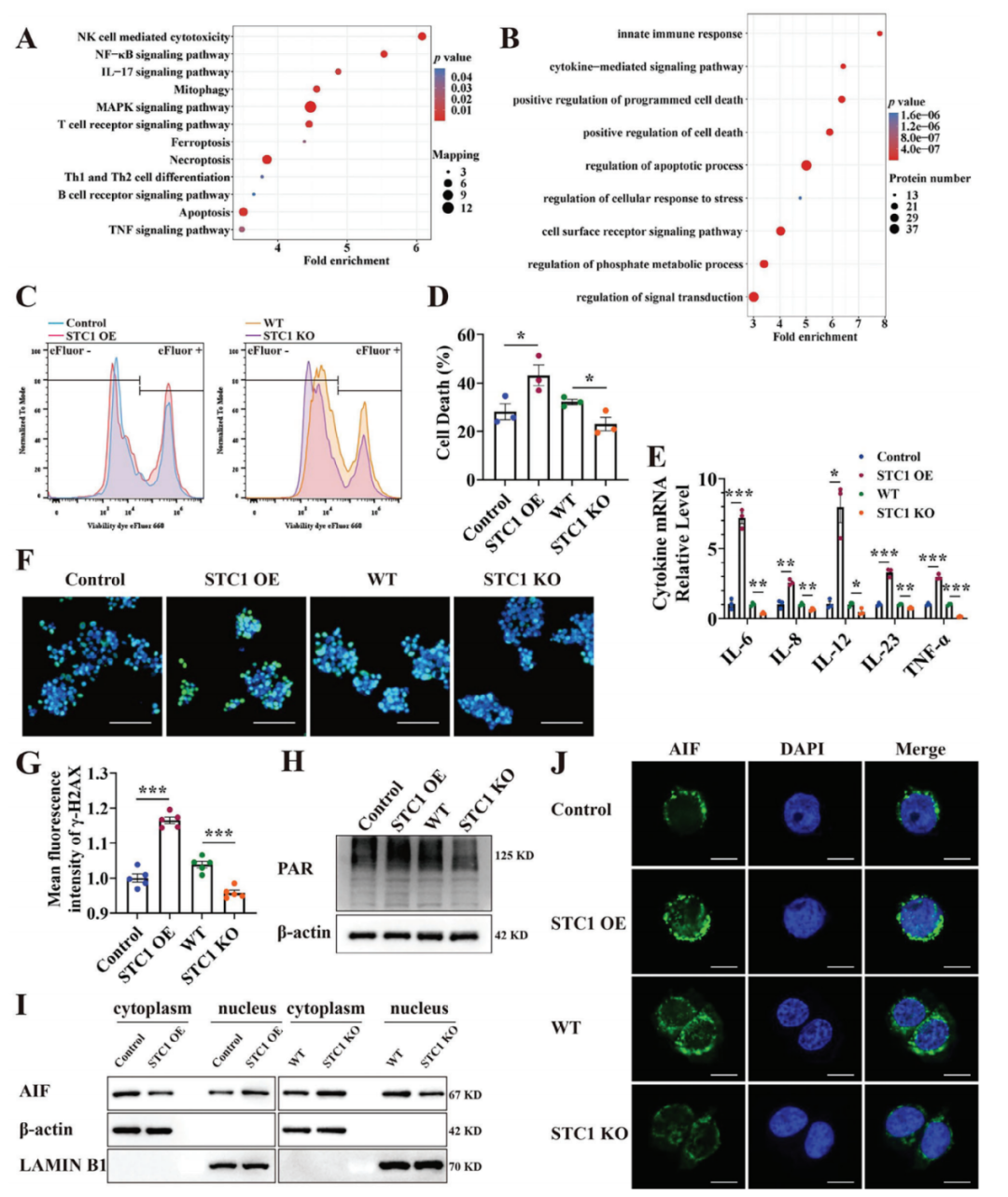
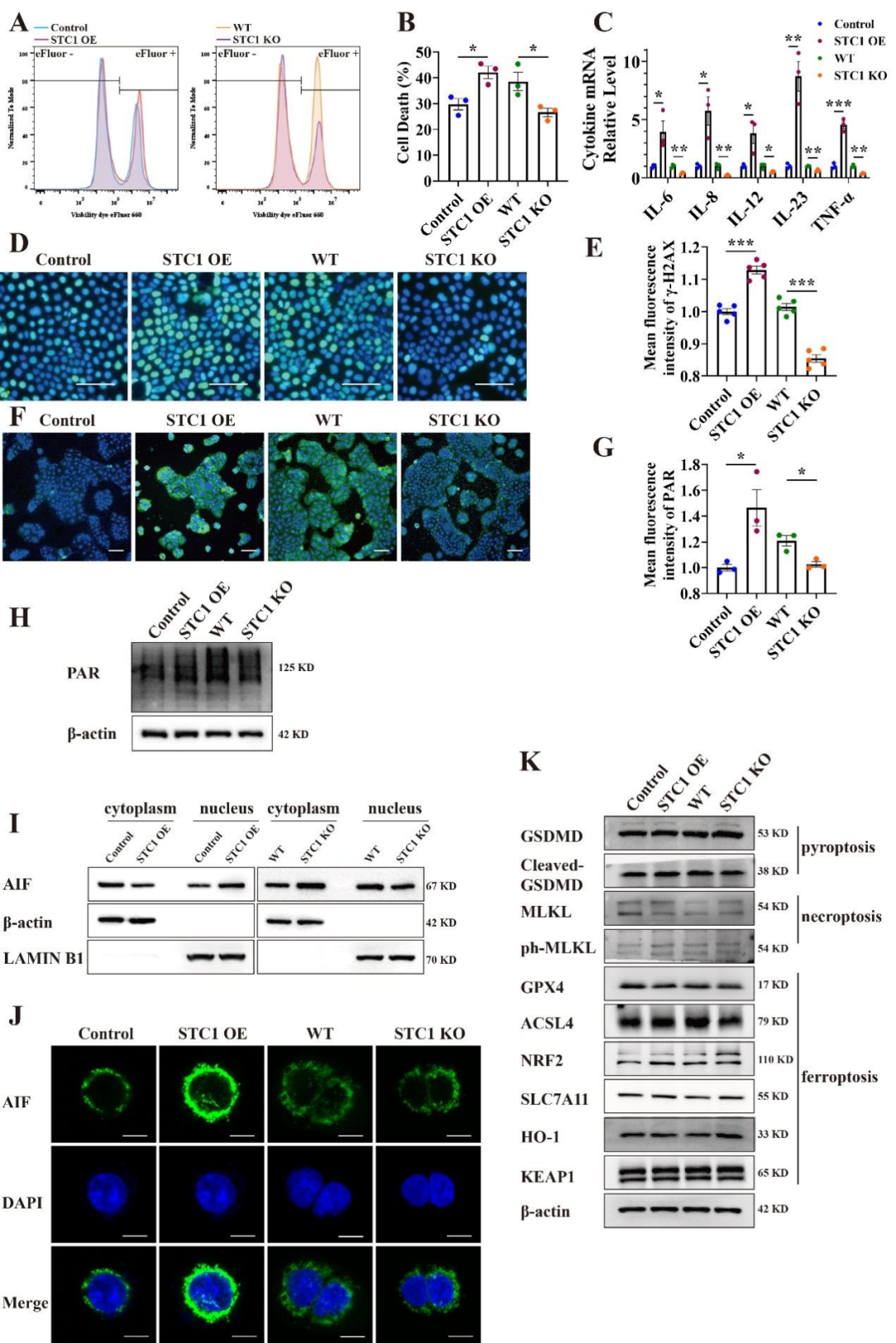
3.STC1在体外加剧PARP1依赖性细胞死亡和炎症反应
在体外实验中,研究人员使用H2O2来诱导氧化应激,结果显示,STC1在H2O2刺激下表达上调[NCM460细胞(人正常结肠上皮细胞);Caco2细胞(人结直肠腺癌细胞)]。为阐明STC1在体外的功能机制,研究者构建了STC1过表达(STC1 OE)和STC1敲除(STC1 KO)细胞系。对STC1 OE NCM460细胞的蛋白组学分析显示,STC1上调与促炎信号相关,包括NF-κB、丝裂原活化蛋白激酶(MAPK)、TNF和IL-17信号通路。此外,在KEGG通路分析中,STC1上调有助于免疫细胞活化(如T细胞和B细胞受体信号、以及Th1/Th2分化)和细胞死亡(如铁死亡、坏死性凋亡和凋亡)相关蛋白的富集。GO分析进一步揭示,在STC1 OE细胞中,包括先天免疫反应、细胞死亡和应激反应在内的生物过程被上调。研究人员使用Dye eFluor 660染料检测经H2O2处理的NCM460和Caco2细胞中的死亡细胞,实验分组为:1)过表达对照(Control);2)STC1 OE;3)敲除对照(WT);4)STC1 KO。结果显示,STC1 OE细胞中死亡细胞比例增加,而在STC1 KO细胞中比例降低。促炎细胞因子,包括IL-6、IL-8、IL-12、IL-23和TNF-α,在H2O2刺激的STC1 OE细胞中增加,在STC1 KO细胞中减少。此外,STC1 OE的NCM460和Caco2细胞中增加的γ-H2AX表明在氧化应激下发生了更多的DNA DSB,而在STC1 KO细胞中观察到较少的γ-H2AX。在H2O2刺激的STC1 OE细胞中,PAR合成也上调,而在STC1 KO细胞中被抑制。同样,凋亡诱导因子(AIF)在STC1 OE细胞中显著地从细胞质转移到细胞核内,而这一过程在STC1 KO细胞中被阻断。作者还通过蛋白质印迹(western blotting)评估了STC1 OE和STC1 KO细胞中细胞焦亡、铁死亡和坏死性凋亡的公认标志物。然而,在氧化应激条件下,这些标志物的表达并未因STC1的过表达或敲除而发生显著改变。
图3.STC1在体外增强PARP1依赖性细胞死亡,并加重炎症反应
图4.STC1在体外增强PARP1依赖性细胞死亡,同时加重氧化应激诱导的炎症反应
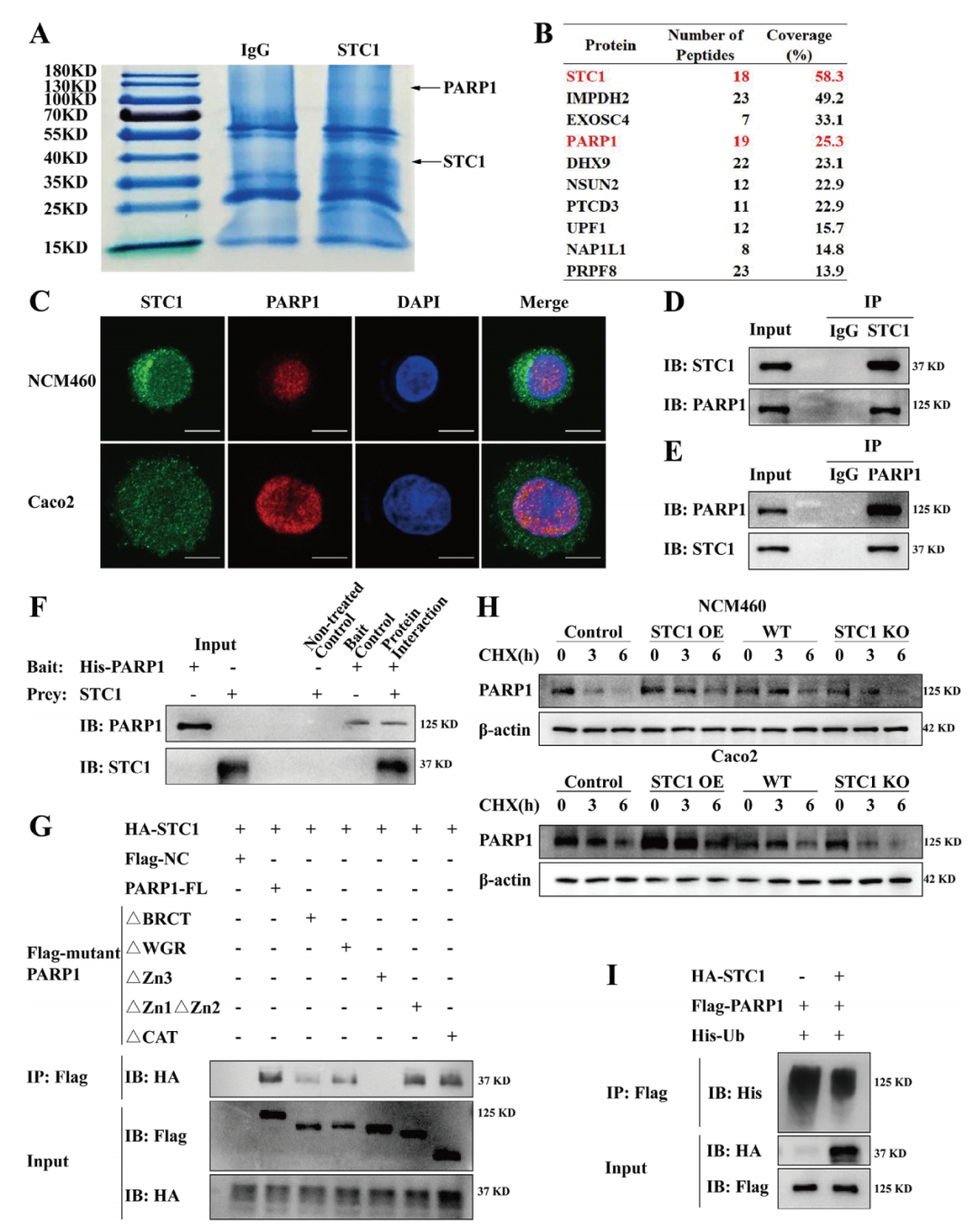
4.在氧化应激诱导的炎症环境中,STC1与PARP1相互作用并上调PARP1,并通过PARP1-JNK互作激活JNK通路
研究人员对STC1 OE NCM460细胞的免疫沉淀产物进行高通量质谱(MS)和蛋白组学分析,以鉴定与STC1相互作用的蛋白。由此研究人员鉴定出PARP1为与STC1蛋白相互作用的潜在候选分子。进一步分析STC1和PARP1蛋白的亚细胞分布发现,在NCM460和Caco2细胞中,STC1与PARP1共定位于细胞核。通过免疫共沉淀(co-IP)和His下拉分析证实了STC1与PARP1的相互作用。此外,作者设计了5种PARP1突变体,蛋白截短分析显示,全长PARP1蛋白以及缺失前两个锌指结构域(△Zn1△Zn2)、自我修饰结构域(△BRCT和△WGR)和催化结构域(△CAT)的截短突变体均能结合STC1蛋白,而△Zn3截短突变体无法结合STC1,这表明PARP1可能通过Zn3结构域与STC1相互作用。
此外,研究人员在CD患者以及DSS和TNBS诱导的急性小鼠结肠炎模型结肠组织中验证了PARP1蛋白的上调。随后作者通过评估STC1是否影响PARP1蛋白稳定性,以探究PARP1蛋白上调的原因。采用环己.酰亚胺(CHX)抑制蛋白合成,结果显示CHX处理后STC1 OE细胞中PARP1蛋白丰度更高,而STC1 KO细胞中PARP1蛋白迅速降解,这表明STC1通过维持PARP1蛋白稳定性导致其上调。此外,在STC1存在时,PARP1的泛素化水平降低,提示STC1通过抑制PARP1的泛素化来维持其蛋白稳定性。
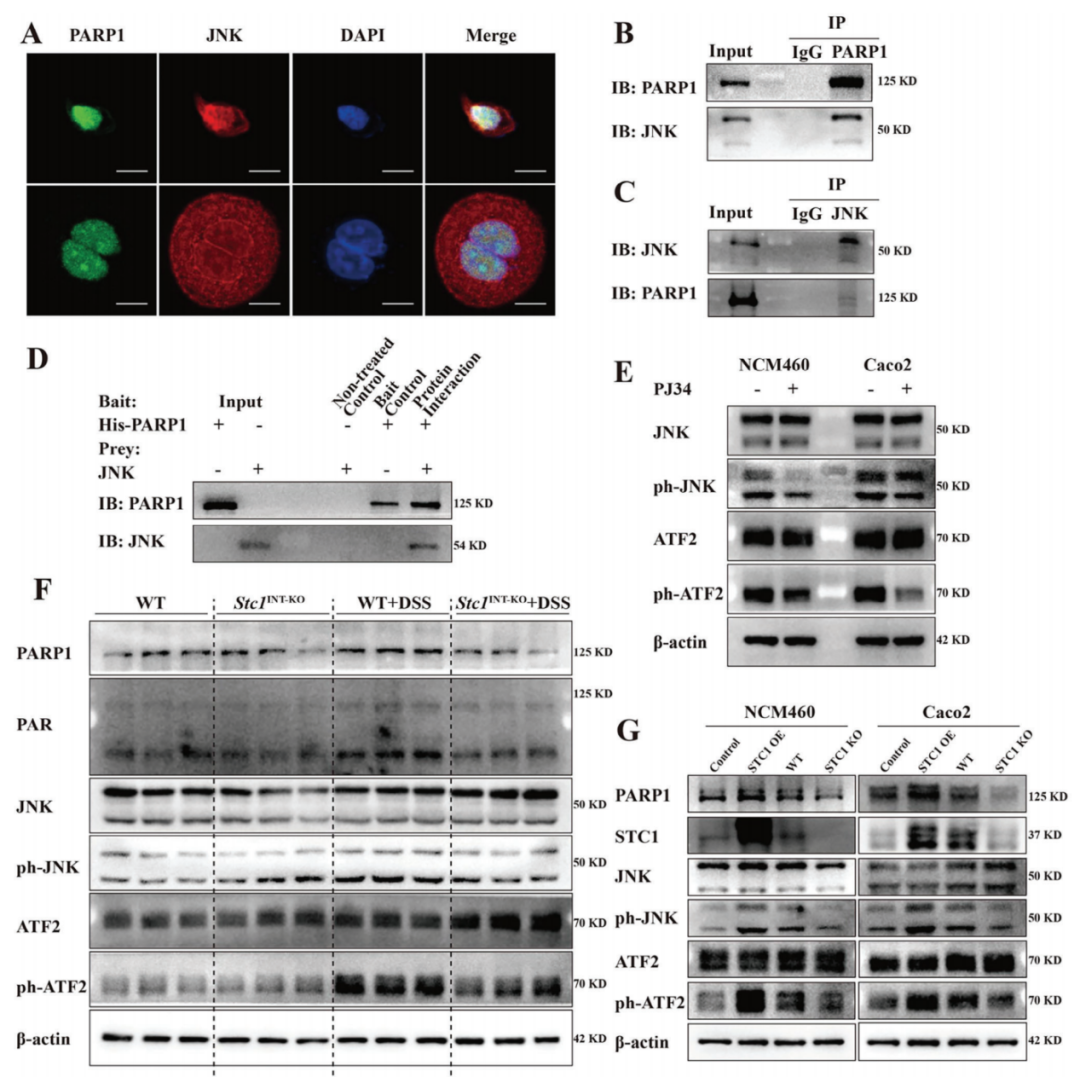
有报道称,PARP1可结合丝裂原活化蛋白激酶8(MAPK8,即JNK)。因此研究者检测了PARP1和JNK蛋白的分布,观察到它们在NCM460和Caco2细胞的细胞核中共定位。通过co-IP和His下拉分析,研究人员也验证了PARP1与JNK的结合。鉴于JNK蛋白是JNK通路的主要效应分子,作者进一步研究了抑制PARP1活性是否会影响JNK通路的激活。用PARP抑制剂PJ34处理细胞后,JNK及其下游效应分子ATF2的磷酸化水平显著下调,表明抑制PARP1会阻碍JNK通路的激活。考虑到STC1 OE细胞中MAPK信号通路显著富集,还评估了STC1是否调控JNK通路。与WT小鼠相比,WT+DSS小鼠中JNK和ATF2蛋白的磷酸化水平上调,而在Stc1INT-KO+DSS小鼠中这一效应被逆转。一致地,在H2O2刺激下,STC1 OE的NCM460和Caco2细胞中PARP1、磷酸化JNK和ATF2的蛋白丰度上调,而STC1 KO细胞中JNK通路的激活受到抑制。这些结果表明,STC1通过与PARP1的相互作用诱导PARP1上调,而PARP1进一步通过与JNK的相互作用调控JNK通路的激活。
图5.STC1与PARP1相互作用并上调PARP1的表达
图6.在氧化应激诱导的炎症环境中,PARP1通过PARP1-JNK相互作用激活JNK通路
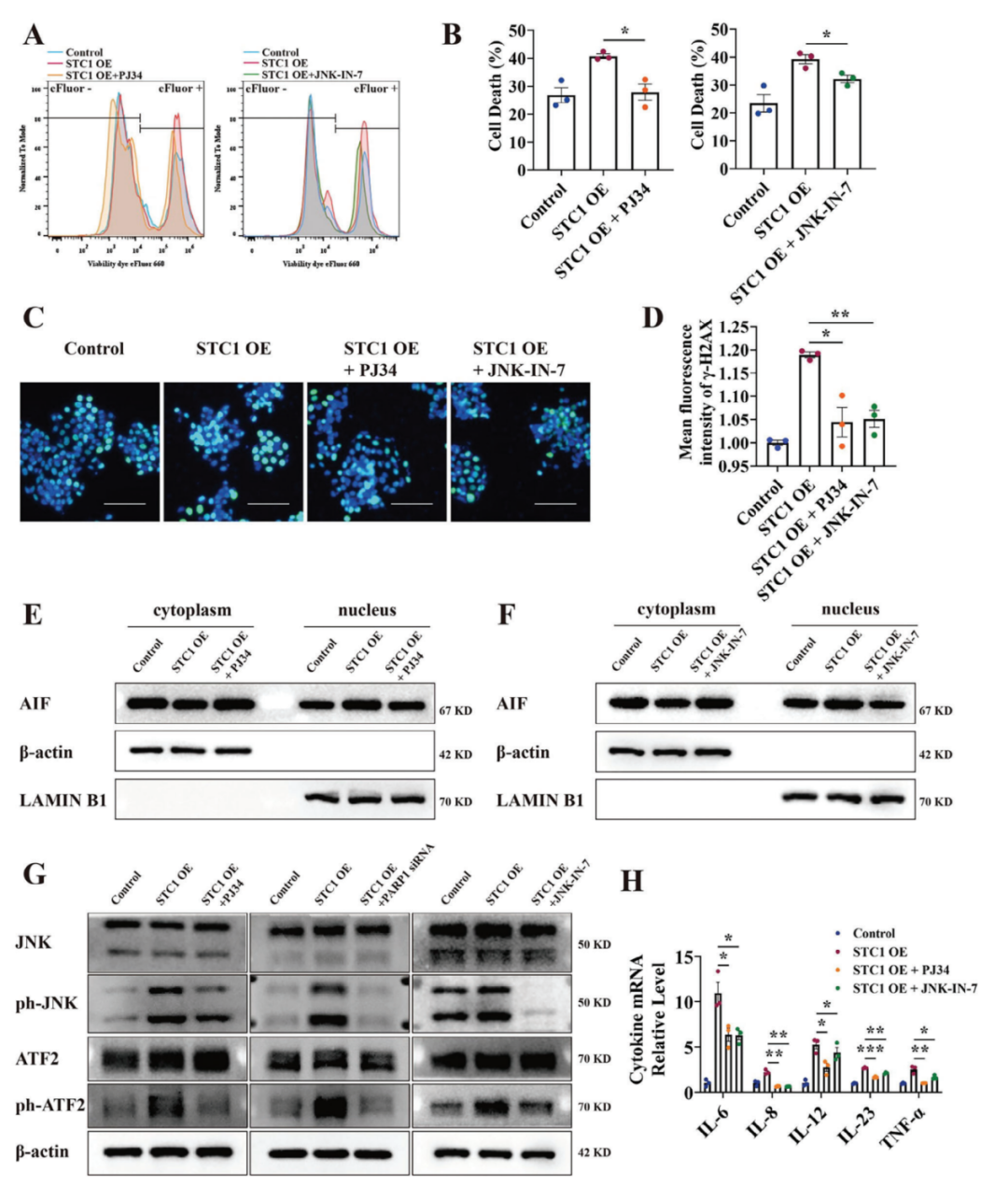
5.抑制PARP1和JNK可减轻PARP1依赖性细胞死亡和氧化应激诱导的炎症反应
接下来,研究人员探究了抑制PARP1和JNK是否能减轻PARP1依赖性细胞死亡和氧化应激相关的炎症反应。在用PARP抑制剂PJ34和JNK抑制剂JNK-IN-7预处理STC1 OE细胞后,H2O2诱导的细胞死亡显著减少,同时伴随DNA DSB频率的降低。PJ34和JNK-IN-7减少了STC1 OE NCM460和Caco2细胞中AIF向细胞核的转位。应用PJ34、PARP1 siRNA和JNK-IN-7在体外也抑制了JNK和ATF2的磷酸化。此外,在H2O2刺激的STC1 OE细胞中,促炎细胞因子,包括IL-6、IL-8、IL-12、IL-23和TNF-α,经PJ34和JNK-IN-7处理后表达下调。
图7.抑制PARP1和JNK可减轻PARP1依赖性细胞死亡和氧化应激诱导的炎症反应
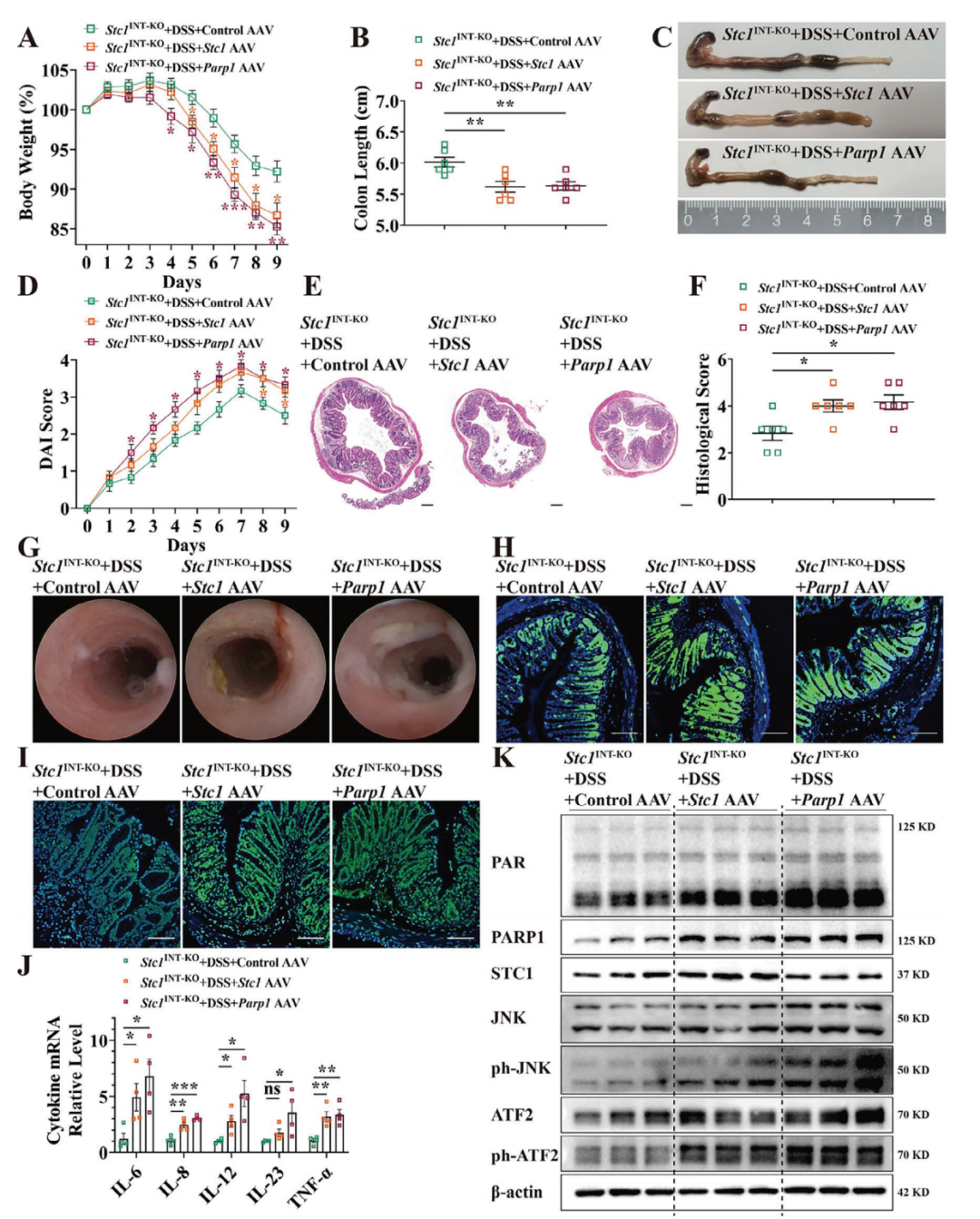
6.恢复Stc1和Parp1表达加重Stc1INT-KO小鼠的DSS诱导性结肠炎
作者进一步研究了恢复Stc1和Parp1表达是否会加重小鼠结肠炎。为恢复Stc1和Parp1的表达,研究者给Stc1INT-KO小鼠腹腔注射腺相关病毒(AAV),并在三周后用DSS诱导结肠炎。在恢复Stc1和Parp1表达后,发生DSS诱导性结肠炎的Stc1INT-KO小鼠表现出体重减轻加剧、结肠长度缩短、DAI评分升高、结肠上皮组织学损伤加重以及组织学评分升高。恢复Stc1和Parp1表达也导致结肠镜检查时结肠黏膜损伤更严重和内镜结肠炎评分更高。此外,PARP1依赖性细胞死亡的指标,包括γ-H2AX和PAR,在Stc1INT-KO小鼠恢复Stc1和Parp1表达后的结肠中均有所增加。TEM显示,在恢复Stc1和Parp1表达后,结肠上皮细胞中线粒体明显肿胀且嵴断裂。另外,在恢复Stc1表达的Stc1INT-KO小鼠结肠组织中,促炎细胞因子IL-6、IL-8、IL-12和TNF-α水平升高;而在恢复Parp1表达的Stc1INT-KO小鼠结肠组织中,这些细胞因子以及IL-23的水平均升高。然而,血清IL-6和TNF-α水平未观察到显著变化。此外,恢复Stc1和Parp1表达还促进了PAR的合成以及JNK和ATF2的磷酸化,表明JNK信号通路被激活。
图8.恢复Stc1和Parp1表达加重DSS诱导的Stc1INT-KO小鼠结肠炎
总结
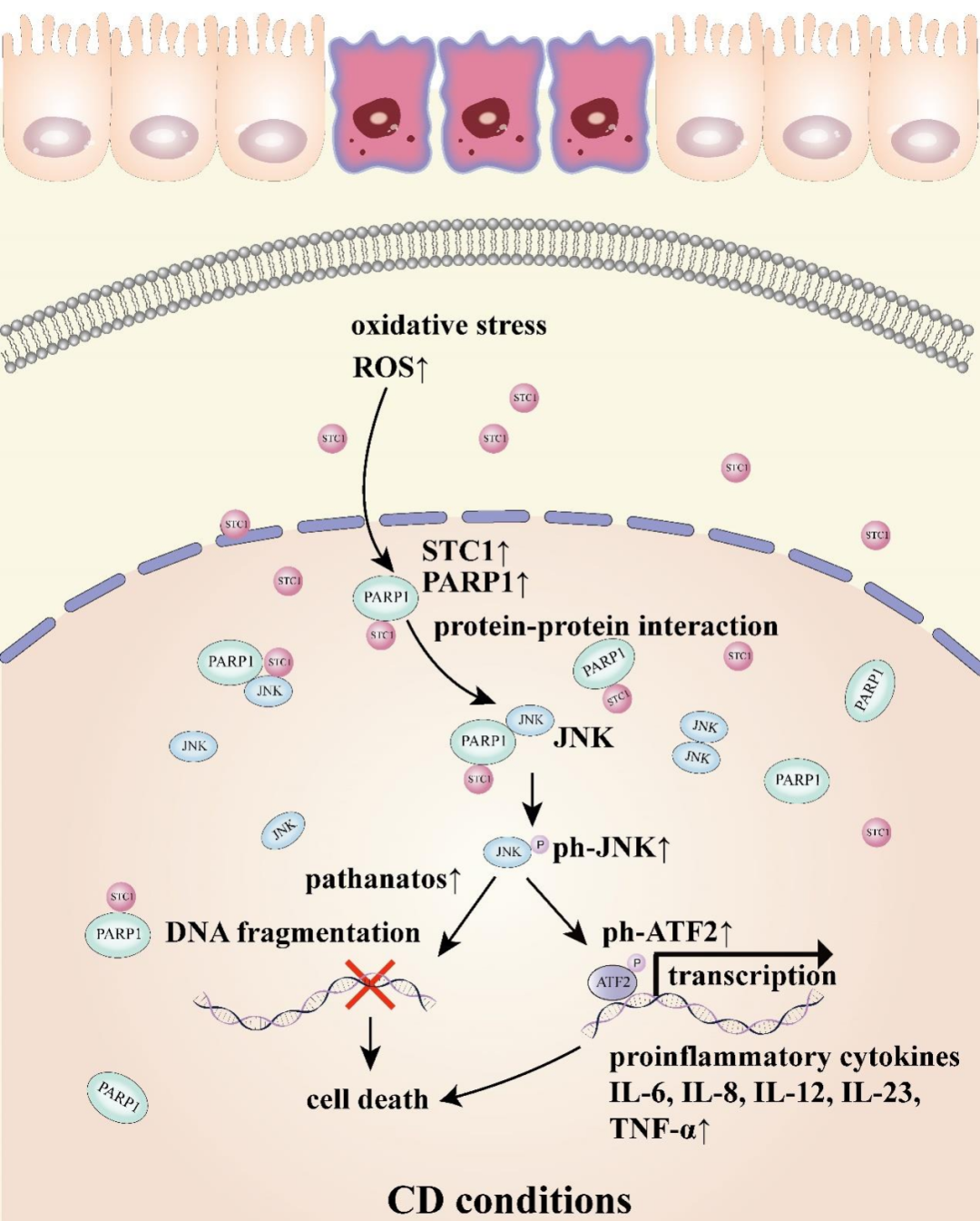
本研究中,STC1的上调加剧了氧化应激下结肠上皮样细胞系的PARP1依赖性细胞死亡并增强了促炎细胞因子的产生,而STC1缺乏则逆转了这些表型。Stc1INT-KO小鼠表现出对DSS诱导的结肠炎的抵抗性,并减轻了PARP1依赖性细胞死亡。此外,作者证实PARP1是STC1的相互作用蛋白,并通过与JNK结合激活JNK通路。同时,抑制PARP1和JNK可逆转由STC1过表达引发的效应。另外,通过恢复Stc1INT-KO小鼠中Stc1和Parp1的表达,研究人员观察到的对DSS诱导结肠炎的抵抗性被消除。因此,STC1通过与PARP1结合,激活JNK通路诱导PARP1依赖性细胞死亡,从而参与CD的发病机制。
综上所述,作者通过STC1-PARP1-JNK轴,阐明了STC1在介导结肠炎中结肠上皮PARP1依赖性细胞死亡和氧化应激相关炎症反应中的重要作用。STC1可能通过诱导PARP1依赖性细胞死亡,在结肠炎的发病机制中扮演重要角色。因此,靶向STC1和PARP1可能是缓解结肠炎的潜在疗法。
STC1-PARP1-JNK通路模式图
本研究得到国家自然科学基金(82270555)和广东省科学技术厅(2021A1515220107)的资助。
通讯作者介绍:
张盛洪,主任医师/教授,消化内科,博士生导师/硕士生导师。自工作以来,一直从事消化内科临床工作,对消化系统疾病(包括肝胆胰疾病)的诊断与治疗有丰富的临床经验,尤其擅长炎症性肠病的临床诊治;熟练掌握胃镜、结肠镜及小肠镜等内镜操作及治疗。研究方向:1、炎症性肠病的临床及基础;2、消化内镜临床研究。以第一作者(含并列第一)和通讯作者发表SCI论文70余篇,多篇论著发表在国际著名刊物如JAMA、Lancet Gastroenterology & Hepatology、Genome Biol、Gut Microbes、Ebiomedicine、Endoscopy、Gastrointest Endosc、J Crohns Colitis、Aliment Pharmacol Ther、Mucosal Immunol、Inflamm Bowel Dis等。获得国自然及省部级基金10余项。
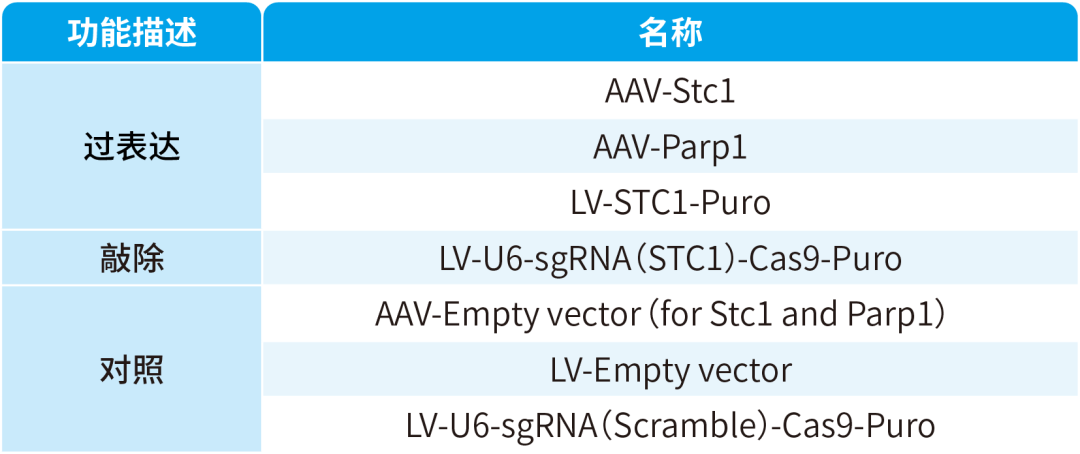
本文使用的病毒产品,列表如下:

了解产品及服务
请扫码添加客服微信:BrainVTA2020

市场:027-65023363 行政/人事:027-62439686 邮箱:marketing@brainvta.com 客服:18140661572(活动咨询、售后反馈等)
销售总监:张经理 18995532642 华东区:陈经理 18013970337 华南区:王经理 13100653525 华中/西区:杨经理 18186518905 华北区:张经理 18893721749
地址:中国武汉东湖高新区光谷七路128号中科开物产业园1号楼
Copyright © 武汉枢密脑科学技术有限公司. All RIGHTS RESERVED.
鄂ICP备2021009124号 DIGITAL BY VTHINK