2025-05-19 阅读量:1252
在终末期肝病(ESLD)的治疗中,肝移植是目前唯—的有效手段,但由于供体器官和细胞的严重短缺,对替代治疗方法的需求变得越来越迫切。此外,肝细胞移植因在高度受损肝脏中植入效果不佳而难以实现临床转化。近年来,通过过表达肝细胞特异性转录因子将成体成纤维细胞直接重编程为肝细胞样细胞的研究,在逆转小鼠肝纤维化方面已显示出积极成果,但如何利用该技术实现具有足够生理功能的肝脏组织再生仍是一个未解决的挑战。肝脏本身是理想的重编程场所,但ESLD患者的受损肝脏环境无法支持原生肝细胞生存,更难以作为细胞重编程和组织再生的场所。异位再生是另一种可行的方法,但由于肝脏体积大且需要复杂的循环系统支持,目前的方法难以实现。因此,利用现有器官(如脾脏)的原生血管网络支持体内肝脏再生成为一种替代方案。将脾脏转化为肝脏具有多种优势:(1)脾脏是一个可有可无的器官(在脾切除手术的临床案例中得到证实),对其操作对身体的影响较小;(2)脾脏空间大、结构松散,适合容纳大量肝细胞;(3)脾脏有丰富的血液供应,可支持再生肝脏的生长和功能化。
南京大学董磊教授、张峻峰教授和澳门大学王春明教授联合在Gut上发表了题为“Reprogramming the spleen into a functioning ‘liver’ in vivo”的研究成果,该研究旨在通过直接重编程脾脏成纤维细胞为肝细胞,将脾脏转化为类似肝脏的器官,以解决肝脏再生的临床难题。研究设计了一个三步程序:首先,通过二氧化硅(SiO₂)颗粒刺激激活脾脏成纤维细胞[指网状成纤维细胞及网状纤维,其特异性标记物:ER-TR7],以产生更多的成纤维细胞和细胞外基质;其次,利用慢病毒(LV)载体表达三种关键转录因子(3TF)——Foxa3、Gata4和Hnf1a,诱导成纤维细胞转化为诱导性肝细胞(iHeps);最后,通过注射肿瘤坏死因子-α(TNF-α),并利用LV介导表皮生长因子(EGF)和肝细胞生长因子(HGF)过表达,进一步扩增iHeps。这些iHeps具有肝脏功能,如糖原储存、脂质积累和药物代谢,并显著提高了90%肝切除小鼠的存活率。本研究通过直接将脾脏转化为类似肝脏的器官,无需细胞或组织移植,成功在小鼠体内建立了基本的肝脏功能,为终末期肝病的治疗提供了潜在的新途径。
1、小鼠脾脏中成纤维细胞的刺激增殖
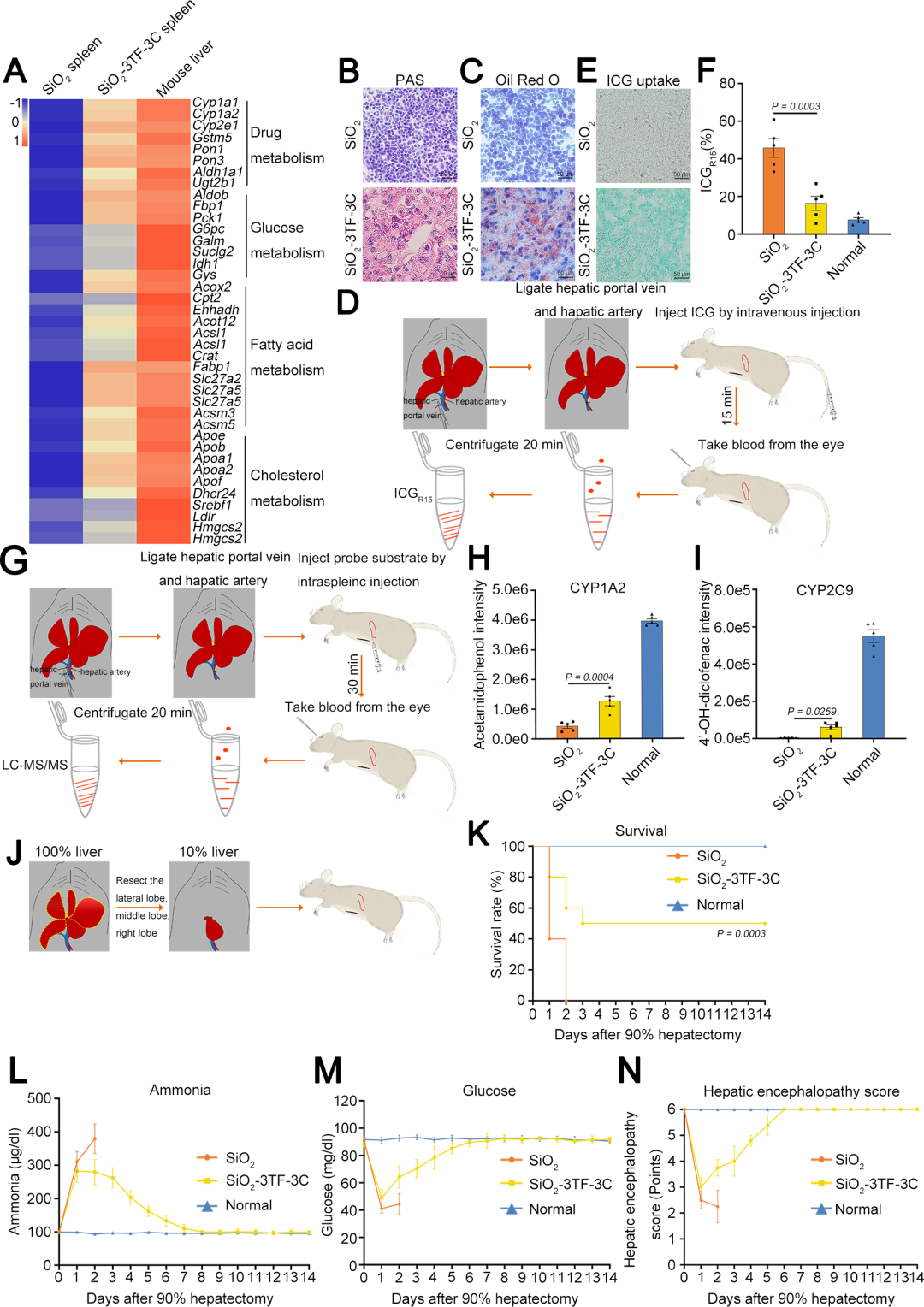
研究人员首先将小鼠脾脏从腹腔内移至皮下位置,这种手术操作不会引起小鼠的生理异常,并且便于通过直接脾内注射的方式反复给药。由于脾脏中成纤维细胞数量较低(不到10%),因此需要在重编程为肝细胞之前增加其数量。团队此前的研究发现,100纳米大小的SiO₂颗粒能够通过激活TGF-β(转化生长因子-β)信号通路有效诱导肺部纤维化,因此团队尝试将其用于诱导脾脏成纤维细胞的增殖。在体内实验中,小鼠每隔4天注射一次SiO₂颗粒(每次1 mg,溶于40 µL PBS),共注射4次,首次注射19天后采集脾脏进行分析。经SiO₂重塑后,脾脏的大小和形态发生了显著变化,脾脏重量平均增加了2.5倍。组织学分析显示,脾脏的白髓和红髓结构显著缩小,而成纤维细胞和纤维化组织明显增加。SiO₂颗粒刺激后,脾脏内ER-TR7⁺网状成纤维细胞显著增殖,EdU(5-乙.炔基-2'-脱氧尿苷)实验(检测细胞增殖)和α-SMA(α-平滑肌肌动蛋白,活化标记物)染色结果均证实了这一点。经过4次SiO₂注射后,脾脏中成纤维细胞的激活遍布整个脾脏,且主要集中在红髓和边缘区。对脾脏成纤维细胞(波形蛋白阳性,Vimentin⁺)的动态计数表明:在第三次脾内SiO₂注射前的两周内,其数量持续增长,但第四次追加注射并未引起进一步增殖。此外,研究人员在第四次注射后继续监测2周,发现成纤维细胞数量逐渐恢复到接近初始水平,表明脾脏的纤维化反应是可逆的。RNA-Seq(RNA测序)分析显示,SiO₂刺激激活了TGF-β1信号通路,这一结果通过qRT-PCR(实时荧光定量逆转录聚合酶链式反应)分析关键基因以及蛋白质印迹(WB)检测p-Smad2(磷酸化Smad2蛋白)和Cyclin D1(细胞周期蛋白D1)表达进一步得到验证。
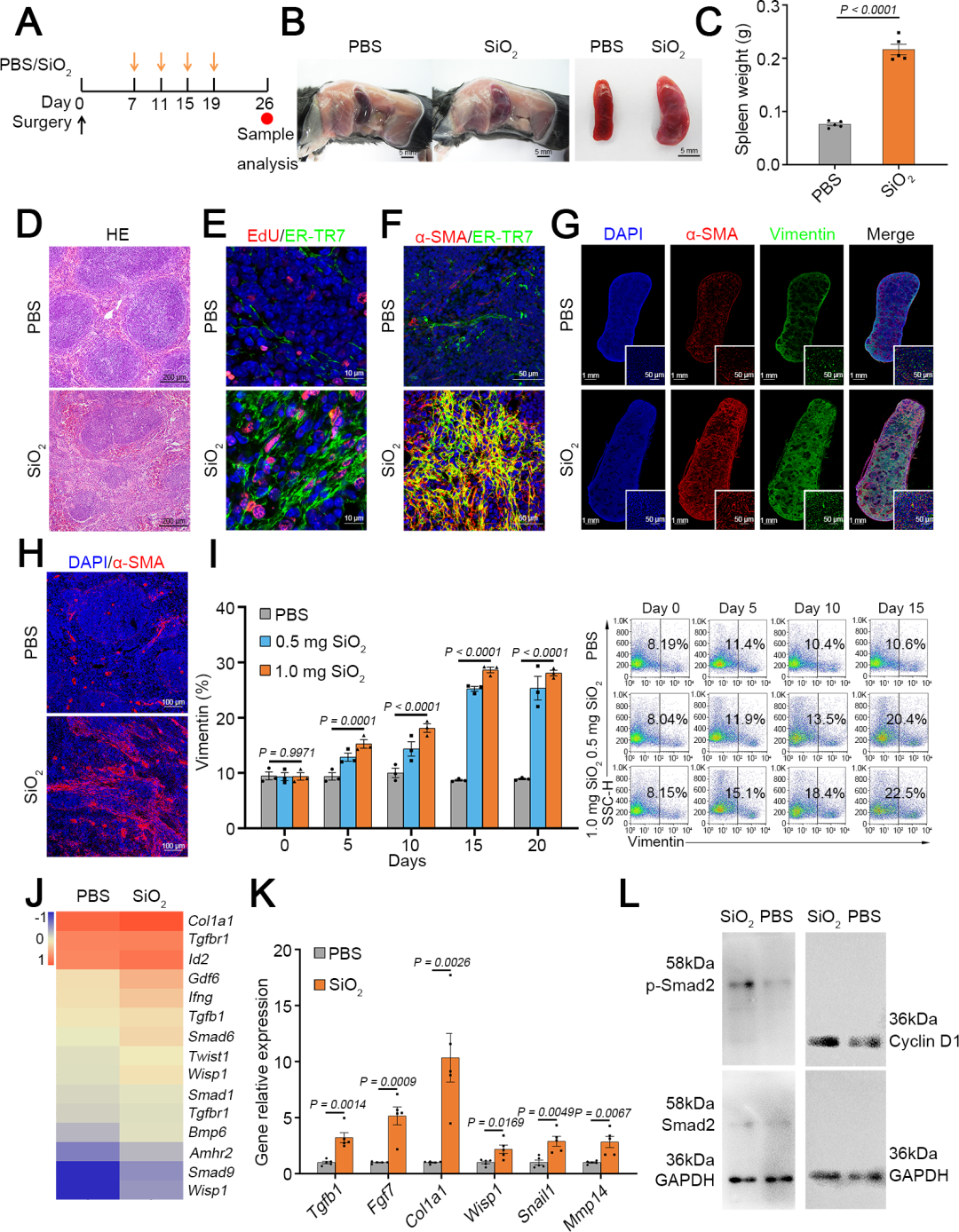
为了评估SiO₂处理的安全性,研究人员在最后一次SiO₂给药后4周分析了小鼠不同组织的情况。结果显示,主要器官的组织学未见明显变化,肝脏也未受到明显损伤,淋巴细胞数量恢复到正常水平。这些结果表明,SiO₂处理不会对动物的长期健康造成损害。综上,SiO₂颗粒处理能够有效增加脾脏中成纤维细胞的数量,且这种增加是可逆的,不会对机体造成长期不良影响。
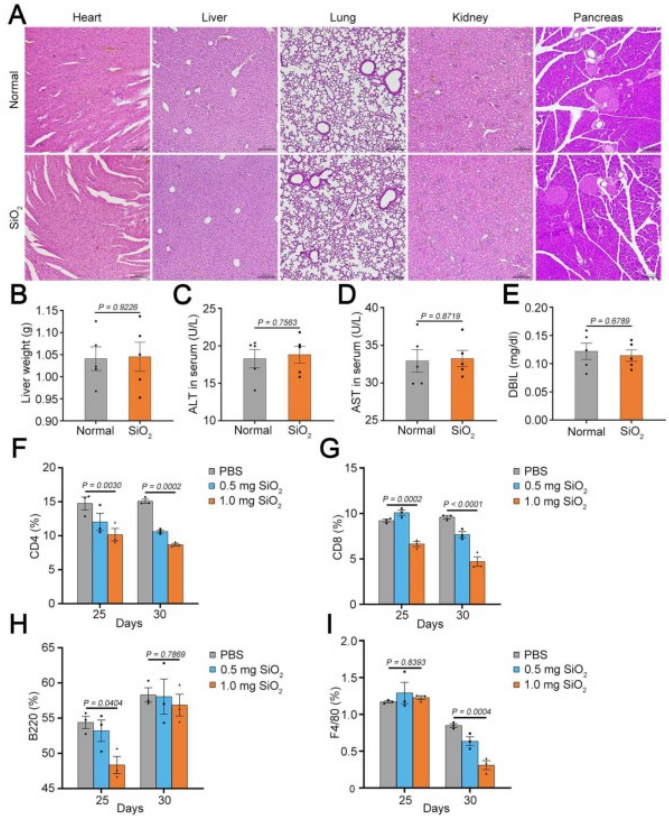
图2. SiO₂处理4周后小鼠不同组织的情况
2、源自SiO₂重塑小鼠脾脏成纤维细胞的诱导性肝细胞的体外表征
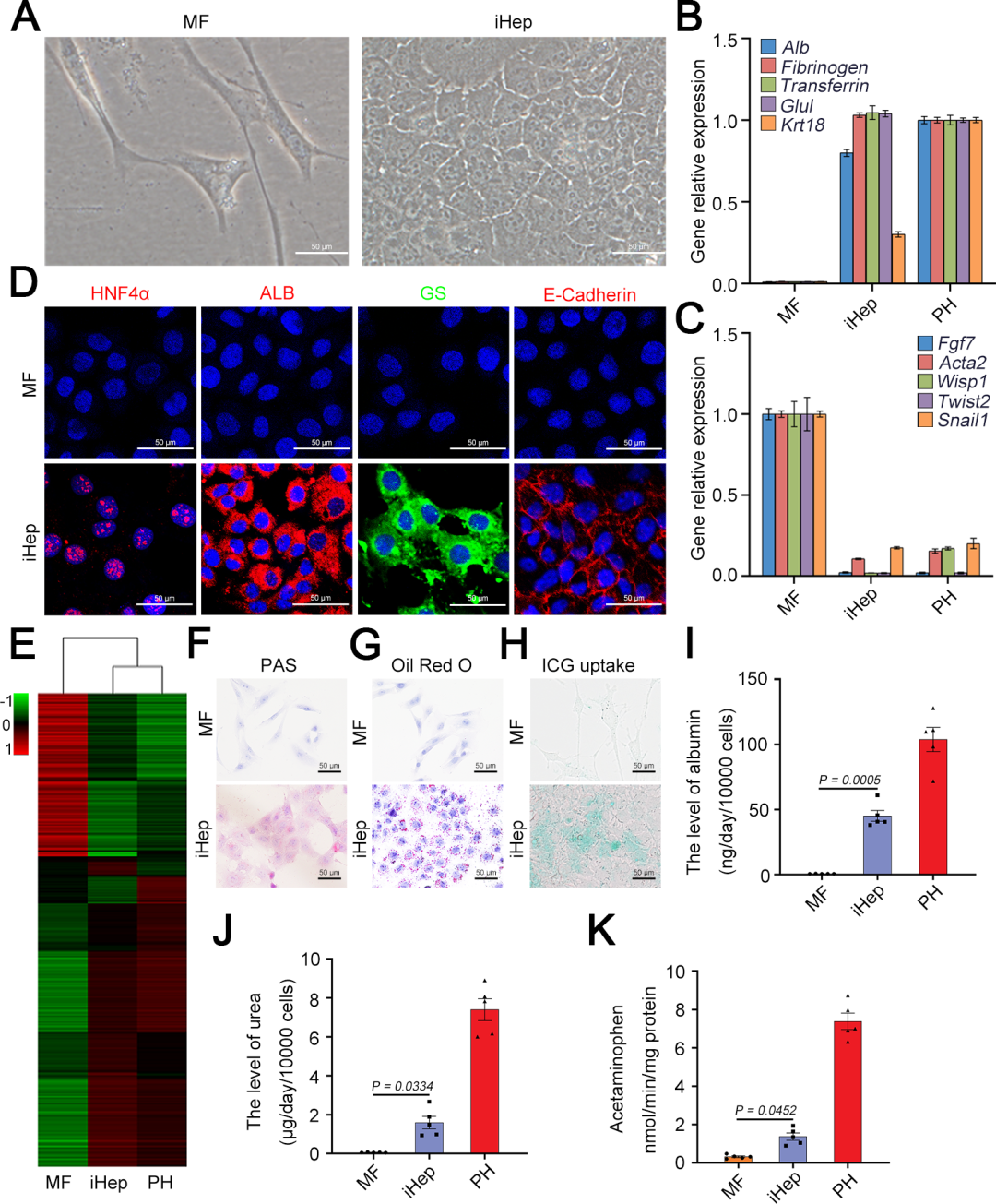
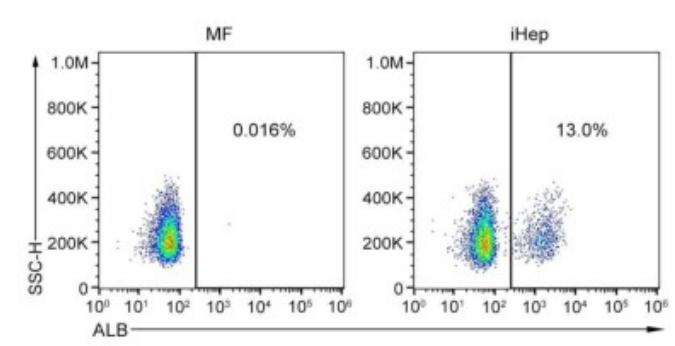
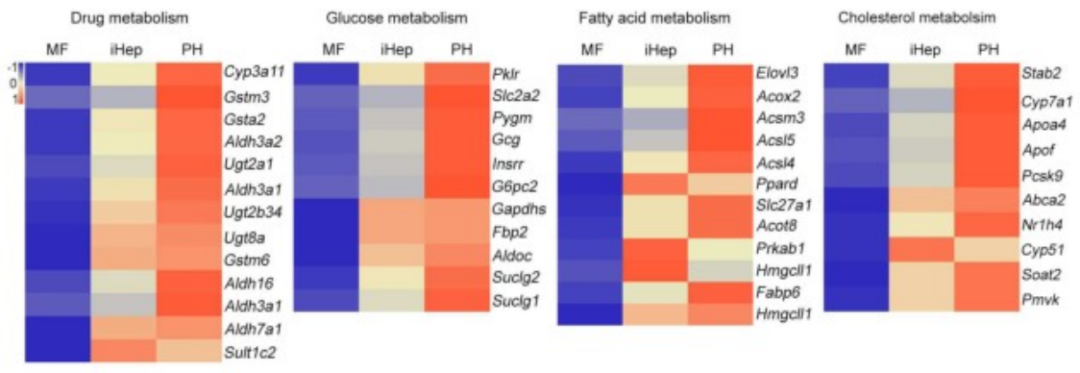
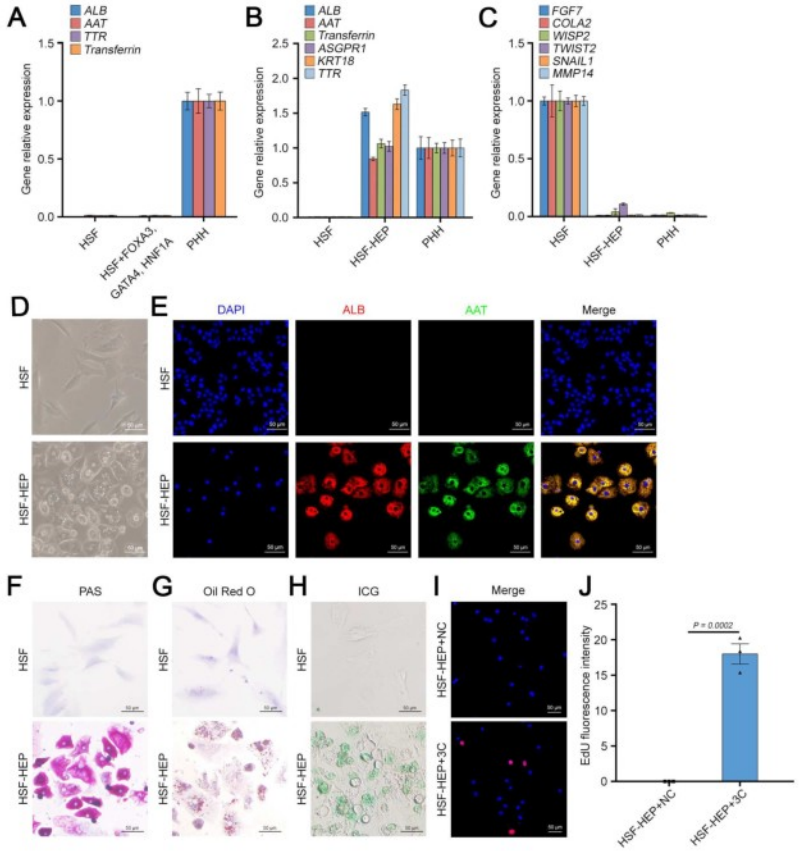
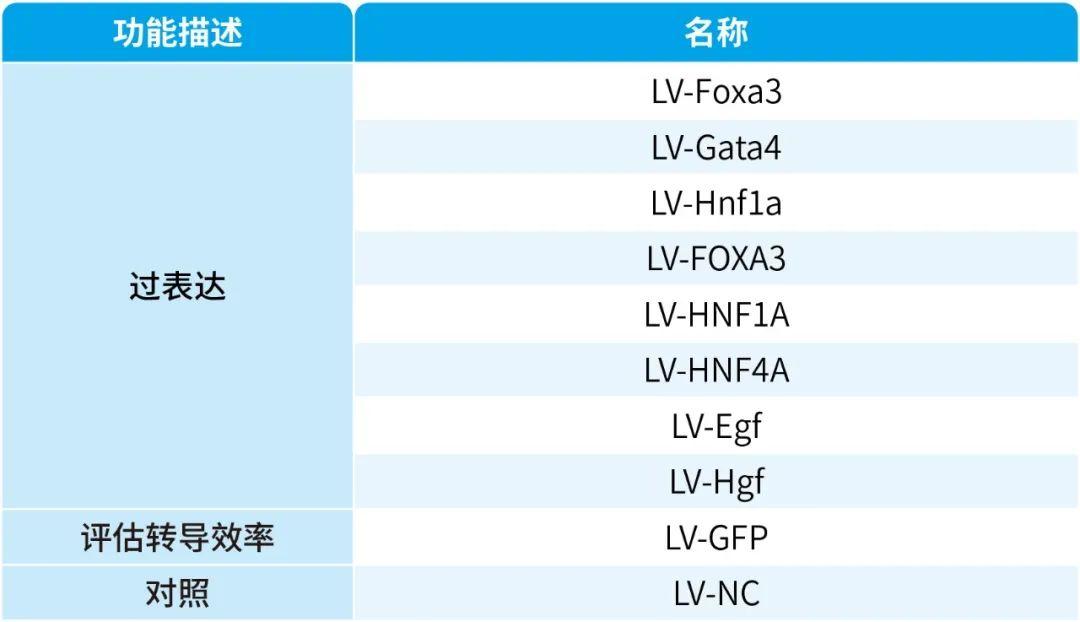
基于之前的研究,研究人员选择了三种关键转录因子(3TF)——Foxa3、Gata4和Hnf1a,将其cDNA分别克隆到LV载体中,以用于实现这些转录因子的过表达。他们首先检测了3TF的过表达是否能在重塑脾脏的成纤维细胞中诱导肝细胞表型。在LV-3TF(LV-Foxa3、LV-Gata4、LV-Hnf1a)转导后的第4天,研究人员通过qRT-PCR证实了小鼠脾脏成纤维细胞(MFs)中3TF的表达。利用LV-3TF体外转导重塑的脾脏成纤维细胞,经过14天的培养后,诱导生成的iHeps表现出典型的上皮细胞形态,并且通过qRT-PCR和RT-PCR(逆转录聚合酶链式反应)分析发现,肝细胞特异性基因在iHeps中显著上调,而成纤维细胞特异性基因则显著下调。免疫荧光染色结果显示,iHeps表达了肝细胞特异性蛋白标志物,包括肝细胞核因子4α(HNF4α)、白蛋白(ALB)、谷氨酰胺合成酶(GS)和E-钙粘蛋白(E-Cadherin)。在转导后第14天,ALB阳性细胞的比例达到13%,表明了有效的肝细胞转分化。此外,微阵列分析显示iHeps的基因表达模式与成年小鼠原代肝细胞(PH)相似,且与MFs有显著差异,特别是在药物代谢以及葡萄糖、脂肪酸和胆固醇代谢相关基因的表达上。研究人员进一步验证了iHeps的典型肝细胞功能。首先,通过PAS(过碘.酸-希夫染色)和油红O染色证实iHeps能够储存糖原和脂肪滴。其次,iHeps能够摄取ICG(吲哚菁绿,反映肝细胞的功能完整性)。此外,iHeps能够分泌ALB和尿素,这是其具有肝脏合成功能的重要标志。iHeps还表现出细胞色素P450(肝细胞药物代谢的核心酶系)的活性,能够将非那西丁代谢为对乙酰氨基酚(APAP)。总之,通过过表达Foxa3、Gata4和Hnf1a,研究人员成功在体外将重塑脾脏成纤维细胞转化为具有成熟肝细胞典型形态和功能特征的类肝细胞。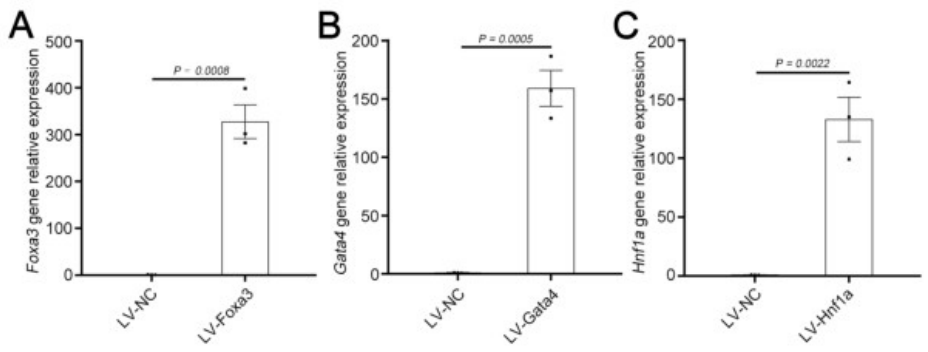
图3. SiO₂重塑小鼠脾脏成纤维细胞来源的iHeps的体外表征
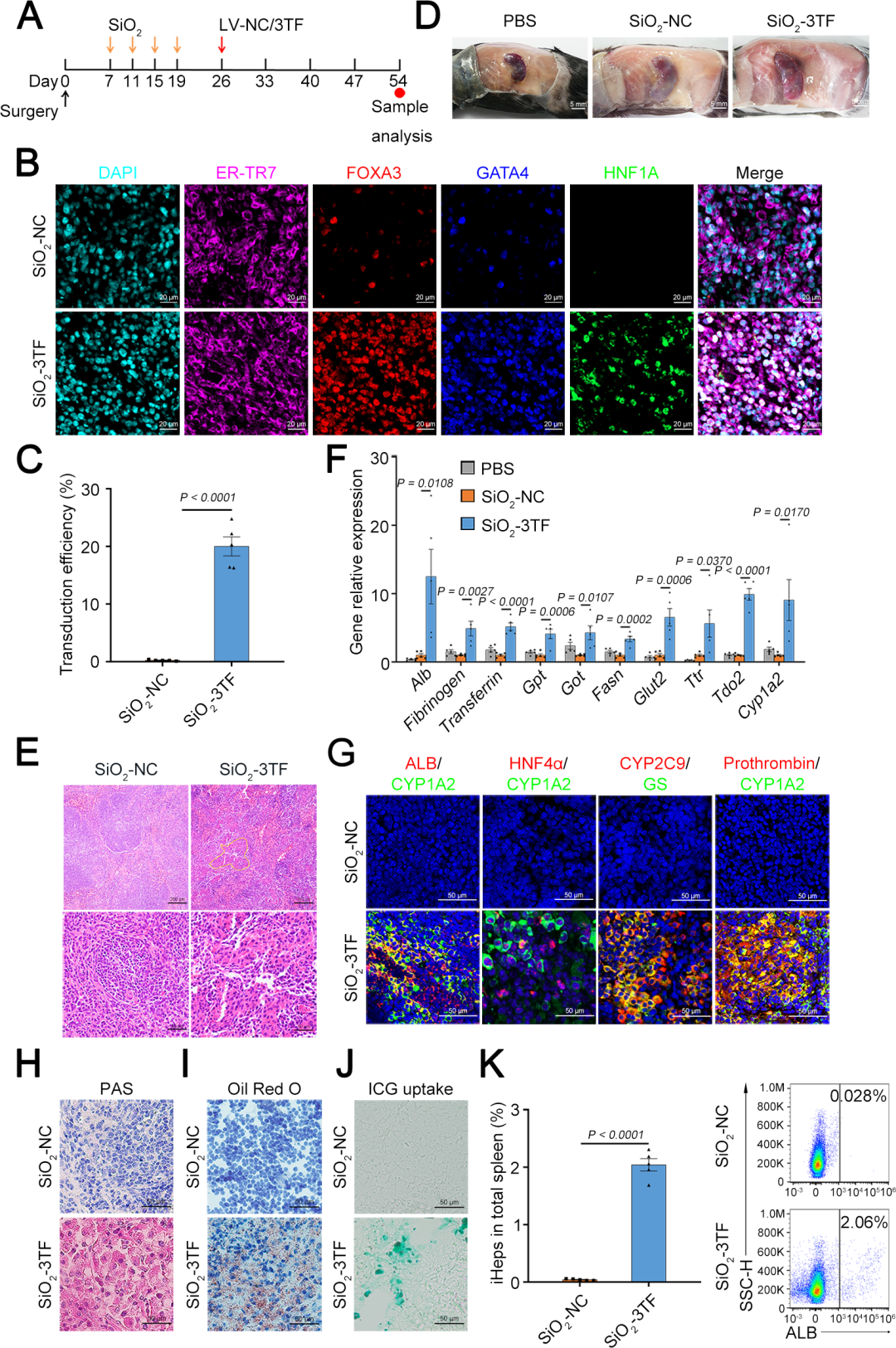
3、体内诱导重塑脾脏成纤维细胞转化为肝细胞
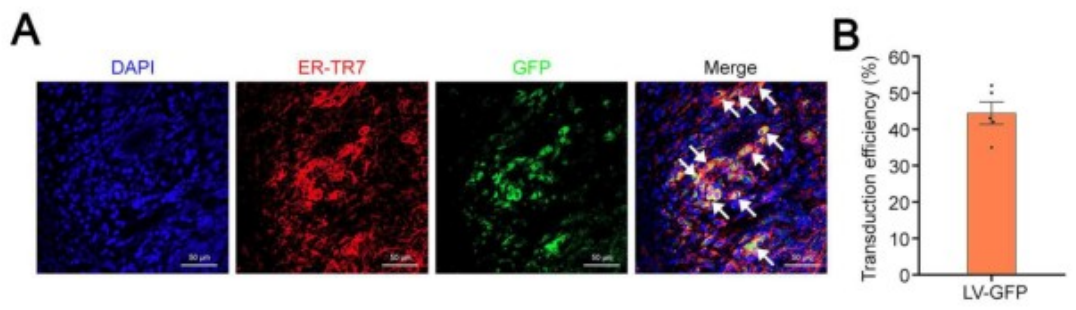
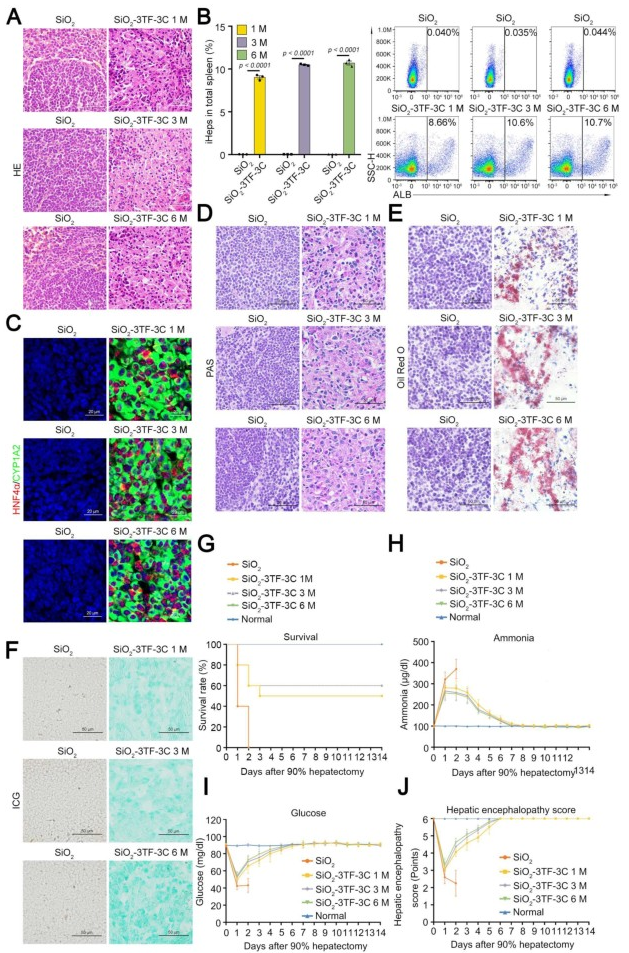
首先,研究人员通过将携带绿色荧光蛋白(GFP)报告基因的慢病毒载体(LV-GFP)注射到经过SiO₂重塑的脾脏中,评估了LV的转导效率。结果显示,LV-GFP能够转导约45%的ER-TR7⁺成纤维细胞。随后,为了检测3TF是否能在体内诱导成纤维细胞转化为iHeps,研究人员将小鼠分为两组:一组接受SiO₂处理后注射携带3TF的慢病毒(LV-3TF),另一组作为对照组(SiO₂-NC组),仅接受空载体(LV-NC)的注射。免疫荧光共染色结果显示,3TF在脾脏成纤维细胞中的转导效率约为20%。在LV-3TF注射后28天,研究人员收集并分析了脾脏组织样本。结果显示,LV-3TF进一步改变了SiO₂重塑脾脏的形态,且在脾脏组织中出现了具有典型上皮细胞形态和多边形结构的iHeps。此外,典型肝细胞特异性基因在LV-3TF处理的脾脏中显著上调。研究人员进一步对脾脏组织进行免疫荧光染色,检测了ALB、细胞色素P450 1A2(CYP1A2)、HNF4α、细胞色素P450 2C9(CYP2C9)、GS和凝血酶原等肝细胞特征标志物,证实了iHeps的存在。更重要的是,LV-3TF处理的脾脏表现出糖原合成、脂质积累和ICG摄取的能力,表明iHeps具备了部分成熟肝细胞的功能特性。最后,研究人员通过流式细胞术定量分析了体内生成的iHeps,发现LV-3TF处理的脾脏中,ALB阳性iHeps占总脾细胞的2.06%。以上结果表明,通过在体内诱导3TF的表达,可以将SiO₂重塑的脾脏成纤维细胞转化为具有肝细胞功能的iHeps。尽管iHeps的总数需要进一步增加以执行真正肝脏的生理功能。
图4. Foxa3、Gata4和Hnf1a在体内诱导SiO₂重塑脾脏成纤维细胞转化为肝细胞
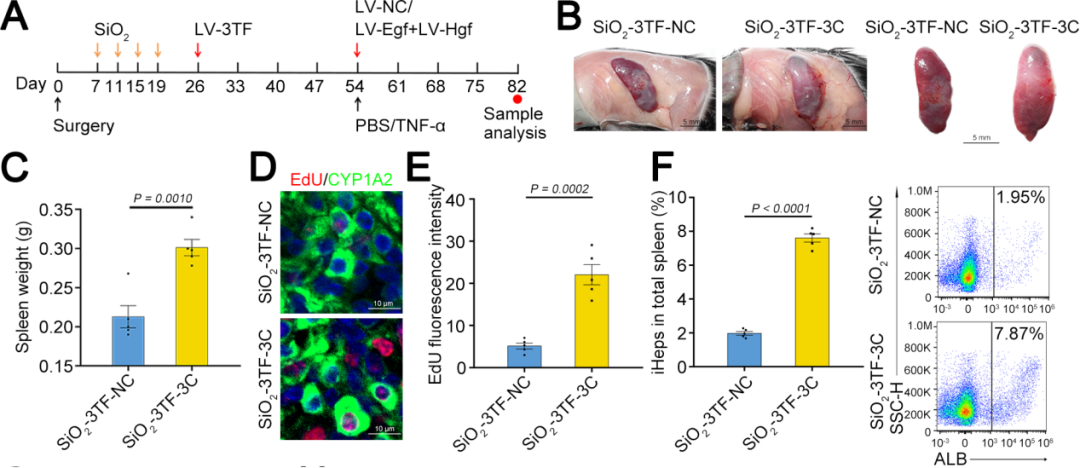
4、iHeps的体内扩增
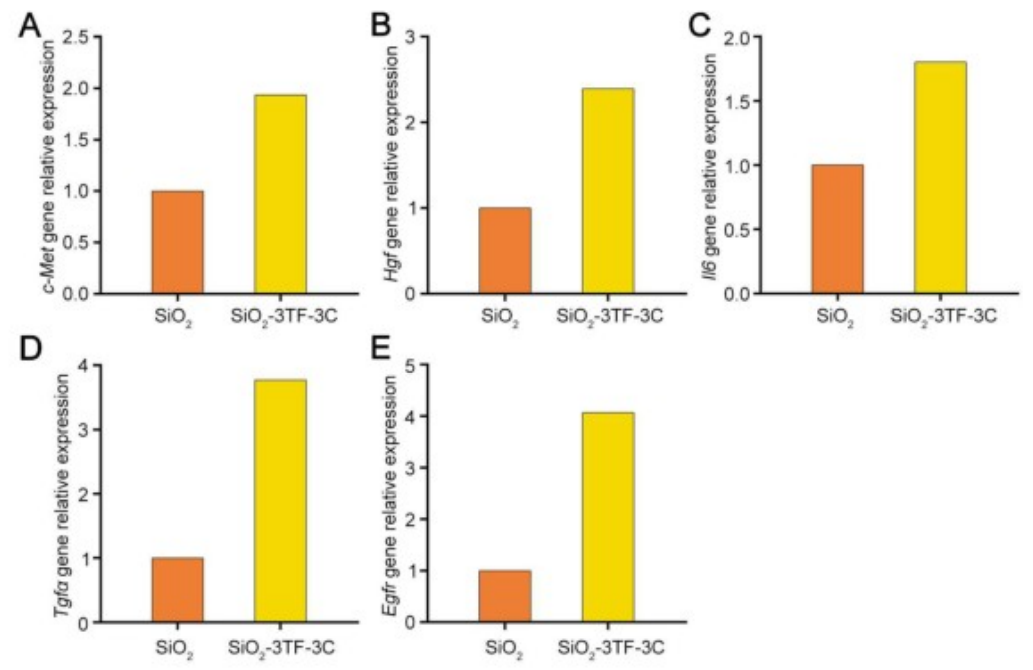
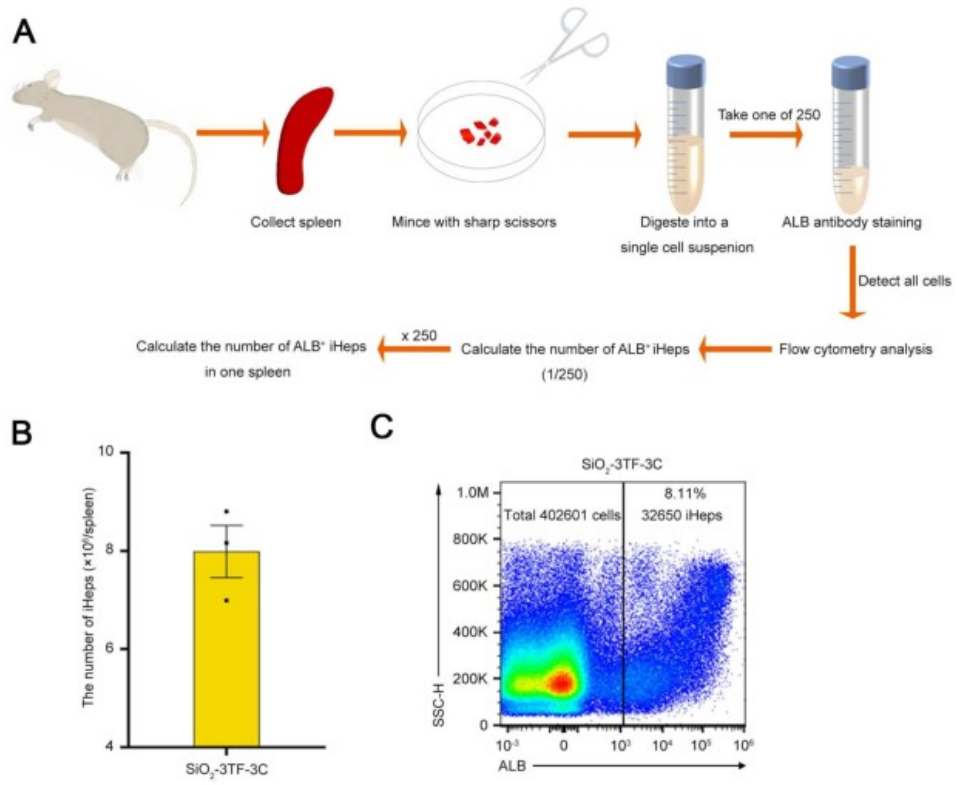
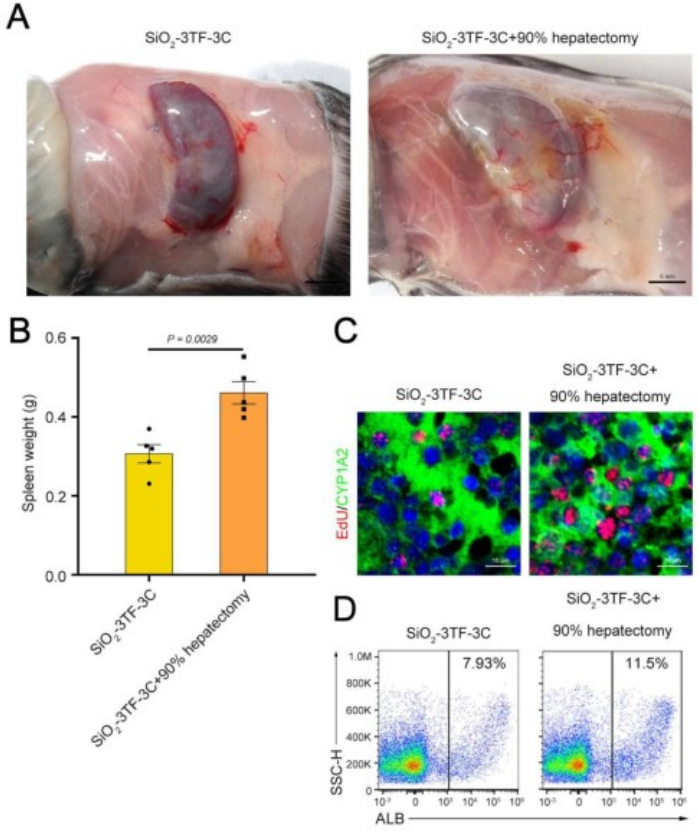
TNF-α是启动肝脏再生和肝细胞增殖的关键因子,能够使肝细胞对其他生长因子产生响应。EGF和HGF是肝细胞的强效有丝分裂原,对肝脏再生至关重要。因此,研究人员通过注射TNF-α蛋白以及利用慢病毒载体(LV-Egf和LV-Hgf)介导的EGF和HGF(简称3C)过表达,来促进3TF处理后的iHeps在脾脏中的增殖,对照小鼠(SiO₂-3TF-NC组)以相同方式处理,但以PBS和空载LV替代3C进行注射。结果显示,经过3C处理的小鼠脾脏(称为“肝化脾”)在形态和重量上发生了显著变化,比对照组小鼠的脾脏更大,暗示iHeps的成功扩增。EdU实验结果显示大量细胞同时检测到CYP1A2和EdU标记,进一步证实了3C刺激后iHeps的增殖,且微阵列分析显示肝化脾中与肝脏再生相关的细胞因子、生长因子及其受体的表达显著上调。此外,肝化脾中iHeps的比例达到7.87%,显著高于仅接受3TF处理的脾脏。荧光激活细胞分选(FACS)分析显示,单个脾脏中的iHeps数量增加到约8×10⁶个,足以执行肝脏的关键生理功能。
图5. iHeps的体内扩增
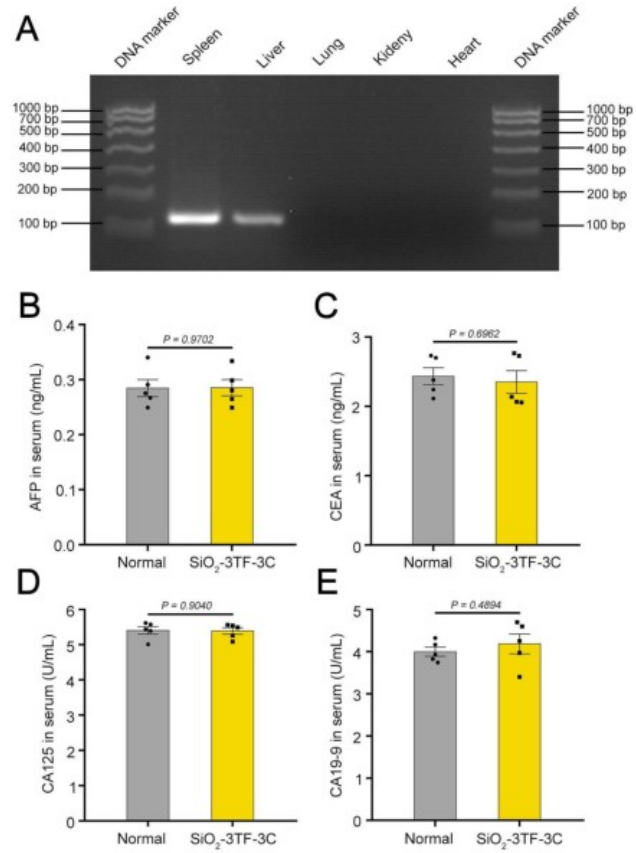
图6. 与肝脏再生相关的细胞因子、生长因子及相关受体的表达情况
图7. 单个脾脏中iHeps数量的测量
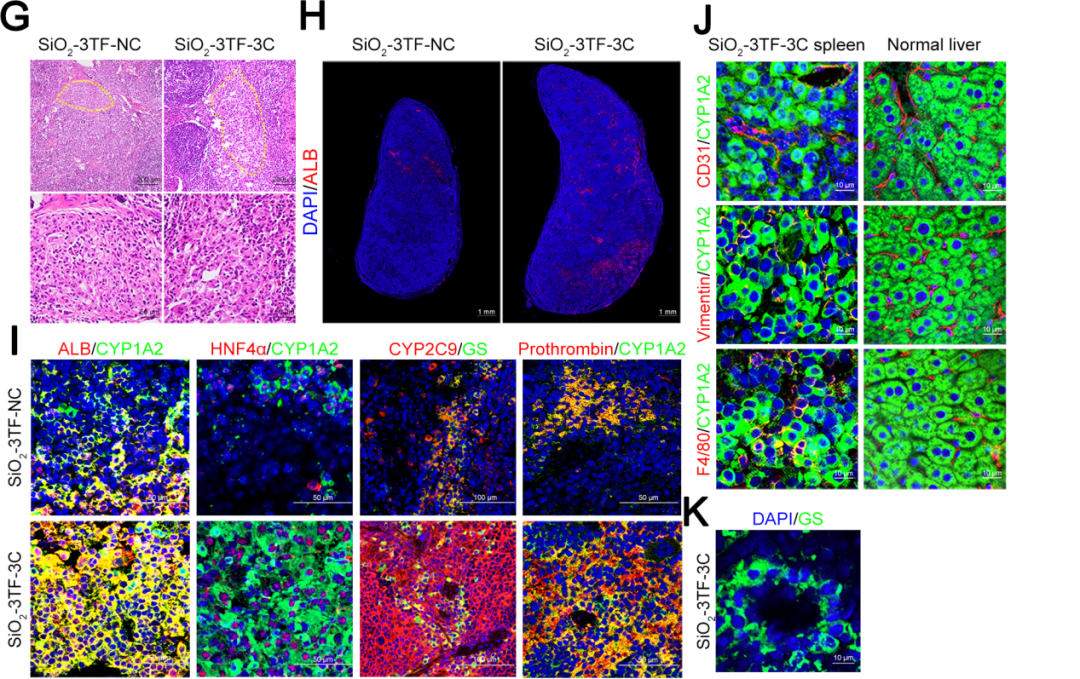
组织学分析显示,3C处理的小鼠脾脏中存在密集排列的多边形细胞,类似于肝细胞,并呈现出类似中央静脉的组织学结构。免疫染色分析揭示了肝化脾中大量表达ALB的iHeps,并证实了ALB、CYP1A2、HNF4α、CYP2C9、GS和凝血酶原等肝细胞标志物的表达。此外,研究人员观察到肝化脾中出现了肝脏微结构,包括血管、成纤维细胞和巨噬细胞,且通过GS免疫染色发现肝化脾中存在类似天然肝脏的分区的结构。为了评估LV载体的生物安全性,研究人员追踪了HIV-1 Ψ(一种LV特异性DNA序列)在不同器官中的分布,发现信号主要集中在脾脏,但很少出现在肝脏。此外,为了评估Egf和Hgf基因持续表达的安全性,研究人员检测了血清中肿瘤标志物[包括甲胎蛋白、癌胚抗原、糖类抗原125(CA125)和糖类抗原19-9(CA19-9)]的表达水平,结果显示肝化脾小鼠未出现显著变化,进一步证实了该方案的安全性。这些结果表明,TNF-α、EGF和HGF的联合应用有效促进了iHeps的增殖,并使其具备了接近真实肝脏的组织结构和功能特征。
图8. 肝化脾组织学特征与实验方案安全性评估
图15. HSFs在体外的转分化及扩增
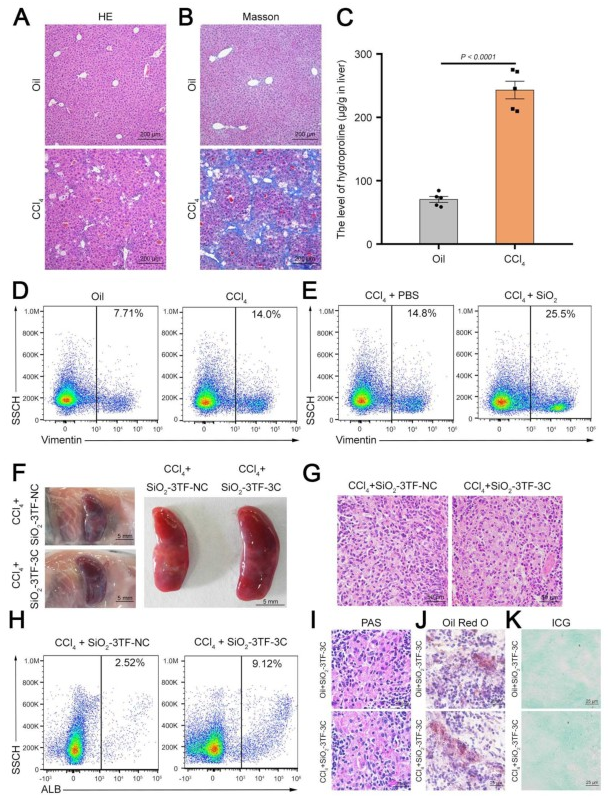
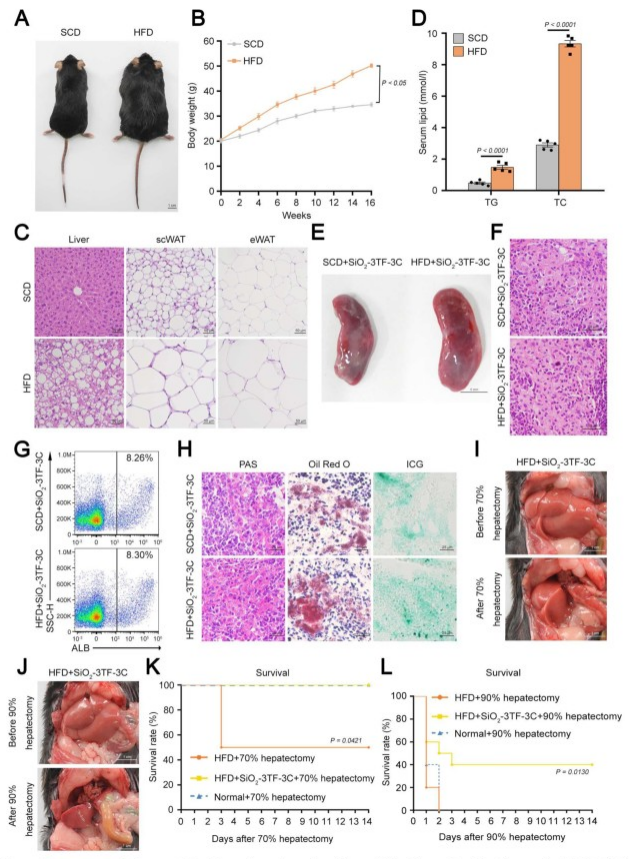
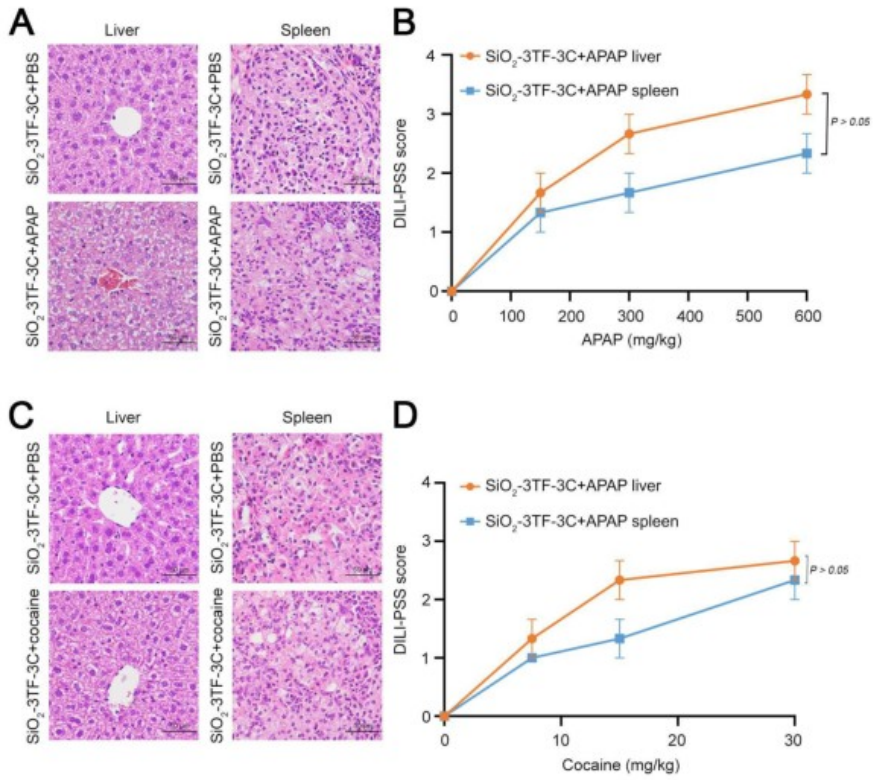
在这项研究中,研究人员通过在小鼠脾脏中原位重编程脾脏成纤维细胞为类肝细胞,成功实现了肝脏功能的体内恢复。与传统的全器官移植、组织工程和细胞移植等手段不同,该策略直接高效地将肝脏功能恢复到能够拯救小鼠受损肝脏的生理水平,且无需移植任何外源细胞或器官,从而避免了免疫排斥和潜在的安全问题,为恢复肝脏功能提供了一种创新且高效的方法,具有潜在的临床应用前景。但仍需进一步的研究以验证其在复杂临床条件下的有效性和安全性。南京大学董磊教授、张峻峰教授和澳门大学王春明教授为本文的共同通讯作者,南京大学刘春艳和王琳涛博士为文章的共同第一作者,本研究得到国家自然科学基金和澳门科学技术发展基金联合科研项目等资助。

通讯作者介绍:
董磊教授,南京大学生命科学学院副院长,无锡市南京大学锡山应用生物技术研究所所长,博士生导师,“长江.学者奖励计划”特聘教授。目前的研究重点是组织与器官的再生,肿瘤的生物治疗,生物材料,药物递送,微生物生长控制等。主要工作发表在Gut、Science Advances、Nature Communications、Cell Reports、eLife等国际权威期刊;获批国家发明专利8项;2项技术成果转化应用;承担重点项目等国家自然科学基金7项,科技部重大新药创制子课题1项;获得教育部新世纪优秀人才、江苏省杰出青年基金、江苏省“六大高峰人才”等人才项目支持。
枢密科技已开发一系列慢病毒现货产品,可提供基因下调、基因过表达、基因敲入/敲除、功能扰动、钙信号检测等现货产品及包装定制服务。质量稳定,周期短!

了解产品及服务
请扫码添加客服微信:BrainVTA2020

市场:027-65023363 行政/人事:027-62439686 邮箱:marketing@brainvta.com 客服:18140661572(活动咨询、售后反馈等)
销售总监:张经理 18995532642 华东区:陈经理 18013970337 华南区:王经理 13100653525 华中/西区:杨经理 18186518905 华北区:张经理 18893721749
地址:中国武汉东湖高新区光谷七路128号中科开物产业园1号楼
Copyright © 武汉枢密脑科学技术有限公司. All RIGHTS RESERVED.
鄂ICP备2021009124号 DIGITAL BY VTHINK