2025-12-15 阅读量:626
肥胖已成为一个全球性的严峻公共卫生挑战,其核心特征是脂肪在脂肪组织中的过度堆积,与代谢性和心血管疾病风险显著增加以及预期寿命缩短密切相关。然而,高效肥胖干预策略的开发仍面临诸多挑战,解析全身能量稳态的核心调控机制,将为制定高效肥胖干预方案提供重要支撑。越来越多的证据表明,中枢神经系统(CNS)在调控全身能量稳态中发挥核心作用。为维持能量稳态,CNS需通过感知并整合外周组织的传入信号,密切监测外周能量状态,进而协调摄食与能量消耗。短期能量平衡调控主要依赖胃肠道信号控制摄食行为,而长期调控则主要依赖脂肪组织信号调节能量储存。
瘦素为脂肪组织分泌的激素,既是能量储存的经典信号,也是中枢能量稳态调控的负反馈调节因子。它主要通过与CNS中表达瘦素受体(LepR)的特定神经元结合,抑制促食欲通路、激活抑食欲神经回路,从而抑制摄食、促进能量消耗。瘦素信号在小鼠、人类等哺乳动物中高度保守,但以中枢对瘦素应答减弱为特征的瘦素抵抗现象,破坏了外周能量储存与中枢的通讯,近30年来一直是抗肥胖药物治疗的主要障碍。尽管已有多种脑内局部机制被提出,但瘦素抵抗的确切分子机制仍不明确。脂肪组织来源的细胞外囊泡(AT-EVs)是脂肪组织与肝脏、骨骼肌、胰岛等器官间通讯的关键介质,在维持全身代谢平衡或引发代谢紊乱中发挥重要作用。然而,AT-EVs在中枢瘦素敏感性调控及能量稳态维持中的作用,仍有待阐明。
2025年11月11日,南京大学医学院附属鼓楼医院毕艳主任医师与南京大学李靓教授团队在国际顶级代谢期刊Cell Metabolism上在线发表了题为“Adipocyte-derived extracellular vesicles are key regulators of central leptin sensitivity and energy homeostasis”的研究成果。该研究证实,脂肪细胞来源的细胞外囊泡(Ad-EVs)是连接脂肪组织与CNS的关键信号载体,通过调节下丘脑回路对瘦素的应答,实现对摄食和体重的核心调控。研究发现,Ad-EVs中携带的一组微小RNA(miRNA)可通过抑制瘦素受体信号通路的负反馈调控因子发挥瘦素增敏作用;而Ad-EVs中这些瘦素增敏性miRNA的缺失,会导致肥胖状态下中枢瘦素抵抗及后续体重增加。更重要的是,研究团队基于Ad-EVs的膜蛋白特征,构建了可靶向CNS的工程化EVs,通过递送瘦素增敏性miRNA,成功逆转了肥胖小鼠的中枢瘦素抵抗并诱导其显著减重。这项工作不仅为理解“脂肪-脑轴”在维持能量平衡中的作用提供了全新视角,更为肥胖的精准治疗提供了极具转化潜力的药物靶点与靶向策略。

1.AT-EVs调控能量平衡
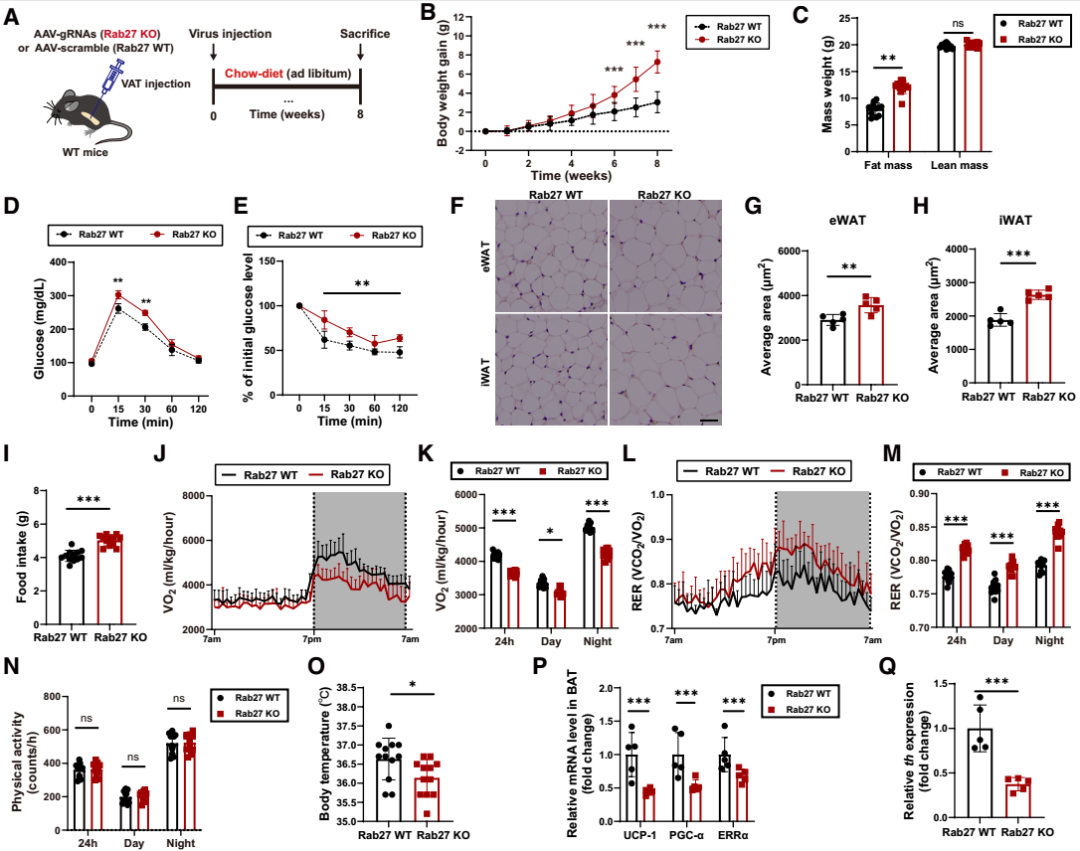
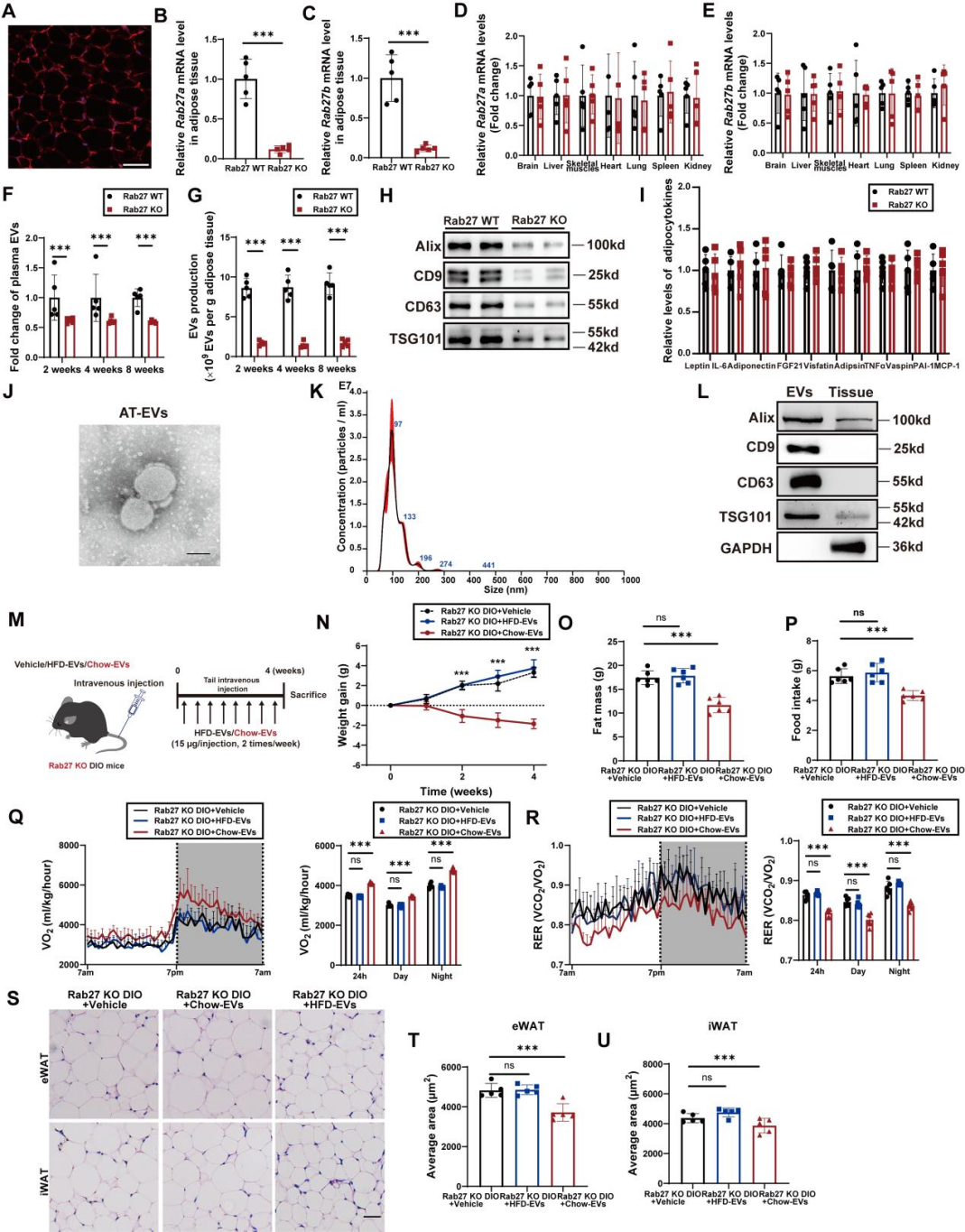
为明确AT-EVs在能量稳态中的作用,研究团队首先构建了脂肪组织特异性EVs生成缺陷小鼠模型:通过向Cas9转基因小鼠的脂肪组织注射携带Rab27(小囊泡分泌至关重要的小GTP酶)向导RNA(gRNA)的腺相关病毒(AAV-gRNA(Rab27)),特异性敲除脂肪组织中的Rab27(Rab27 KO小鼠);对照组小鼠则注射不含gRNA的AAV(Rab27 WT小鼠)。实验验证显示,与Rab27 WT小鼠相比,Rab27 KO小鼠仅在脂肪组织中出现Rab27a/b mRNA水平下降,肝脏、大脑等其他器官无明显变化;纳米颗粒分析与蛋白质印迹(检测EVs标志物CD63、CD9、Alix和TSG101)进一步证实,Rab27 KO小鼠的AT-EVs生成量显著降低,但瘦素、脂联素或成纤维细胞生长因子21(FGF21)等脂肪因子的基因表达未受影响。
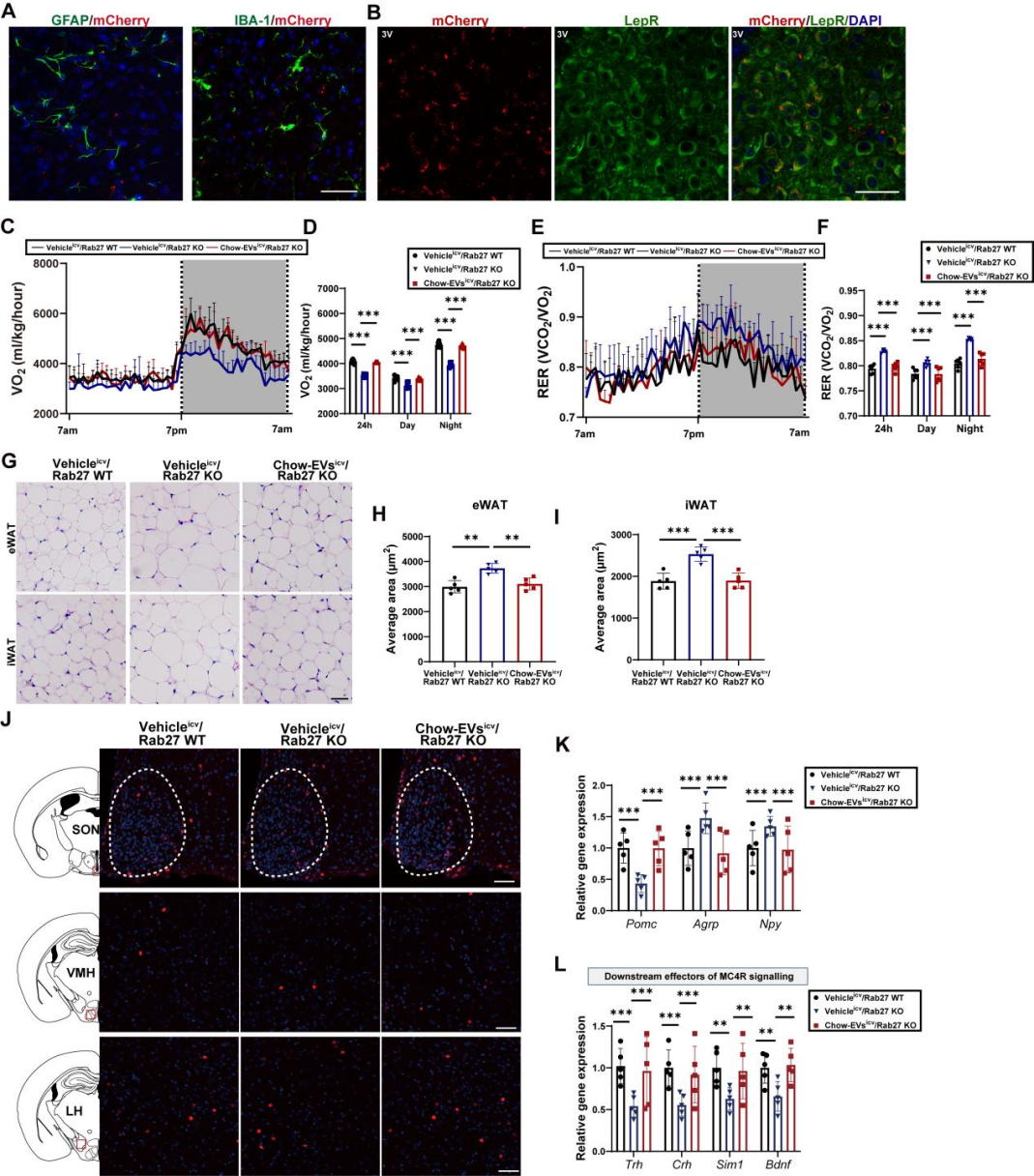
在普通饲料(chow)喂养条件下,与Rab27 WT小鼠相比,Rab27 KO小鼠的体重与脂肪量显著增加,而瘦肉量无差异;葡萄糖耐量试验(GTT)与胰岛素耐量试验(ITT)显示,Rab27 KO小鼠的葡萄糖耐量与胰岛素敏感性显著受损;组织学检查显示,Rab27 KO小鼠的附睾白色脂肪组织(eWAT)与腹股沟白色脂肪组织(iWAT)中的脂肪细胞均大于Rab27 WT小鼠。此外,团队进一步通过代谢笼检测发现:与Rab27 WT小鼠相比,Rab27 KO小鼠的摄食量升高,而耗氧量降低;呼吸交换率(RER)升高(提示其脂肪作为能量来源的利用率较低),而体力活动无显著差异;体温下降,棕色脂肪组织(BAT)中产热基因[如解偶联蛋白1(UCP-1)、雌激素相关受体α(ERRα)等]的表达显著降低,交感神经系统活性标志物酪氨酸羟化酶的水平也显著降低。这些结果表明,AT-EVs的缺失会导致小鼠出现一系列的能量失衡表型。
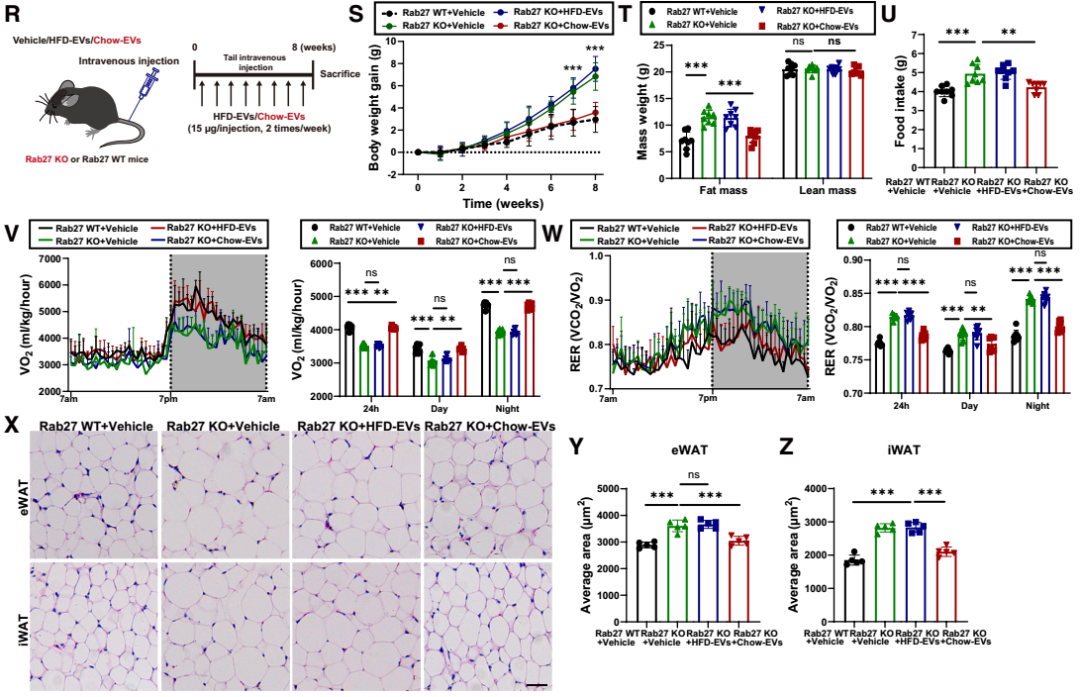
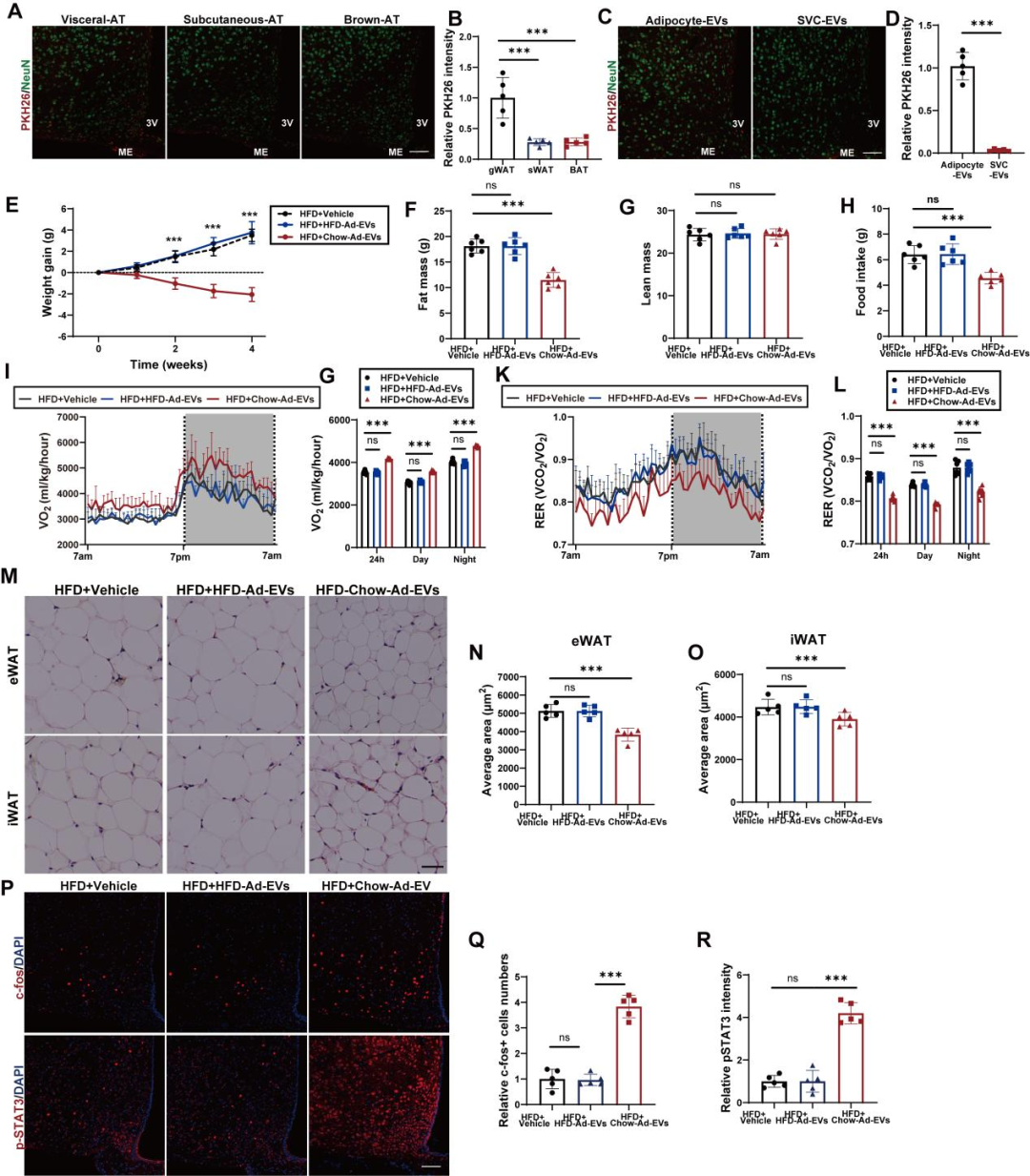
为验证AT-EVs分泌受损是否是导致Rab27缺陷诱导能量失衡的直接原因,研究团队接下来向Rab27 KO小鼠静脉注射不同来源的AT-EVs:从健康chow喂养小鼠提取的“chow-EVs”,或从肥胖高脂饮食(HFD)喂养小鼠提取的“HFD-EVs”。结果显示,与对照组Rab27 KO小鼠[注射空递送载体(vehicle)]相比,chow-EVs组小鼠体重降低、日摄食量减少、能量消耗增加、RER下降,脂肪细胞体积缩小;而HFD-EVs的处理对上述指标无明显影响。为严格评估肥胖状态下AT-EVs的功能,研究团队还在“饮食诱导肥胖(DIO)小鼠”中重复了上述实验:向DIO Cas9转基因小鼠的脂肪组织注射AAV-gRNA(Rab27)从而构建“Rab27 KO DIO小鼠模型”,随后静脉注射chow-EVs或HFD-EVs,结果与前面一致——仅chow-EVs可改善Rab27 KO DIO小鼠的能量代谢异常。这些结果表明,健康的AT-EVs在维持能量稳态中发挥关键作用。
图1. 特异性抑制AT-EVs导致能量平衡受损
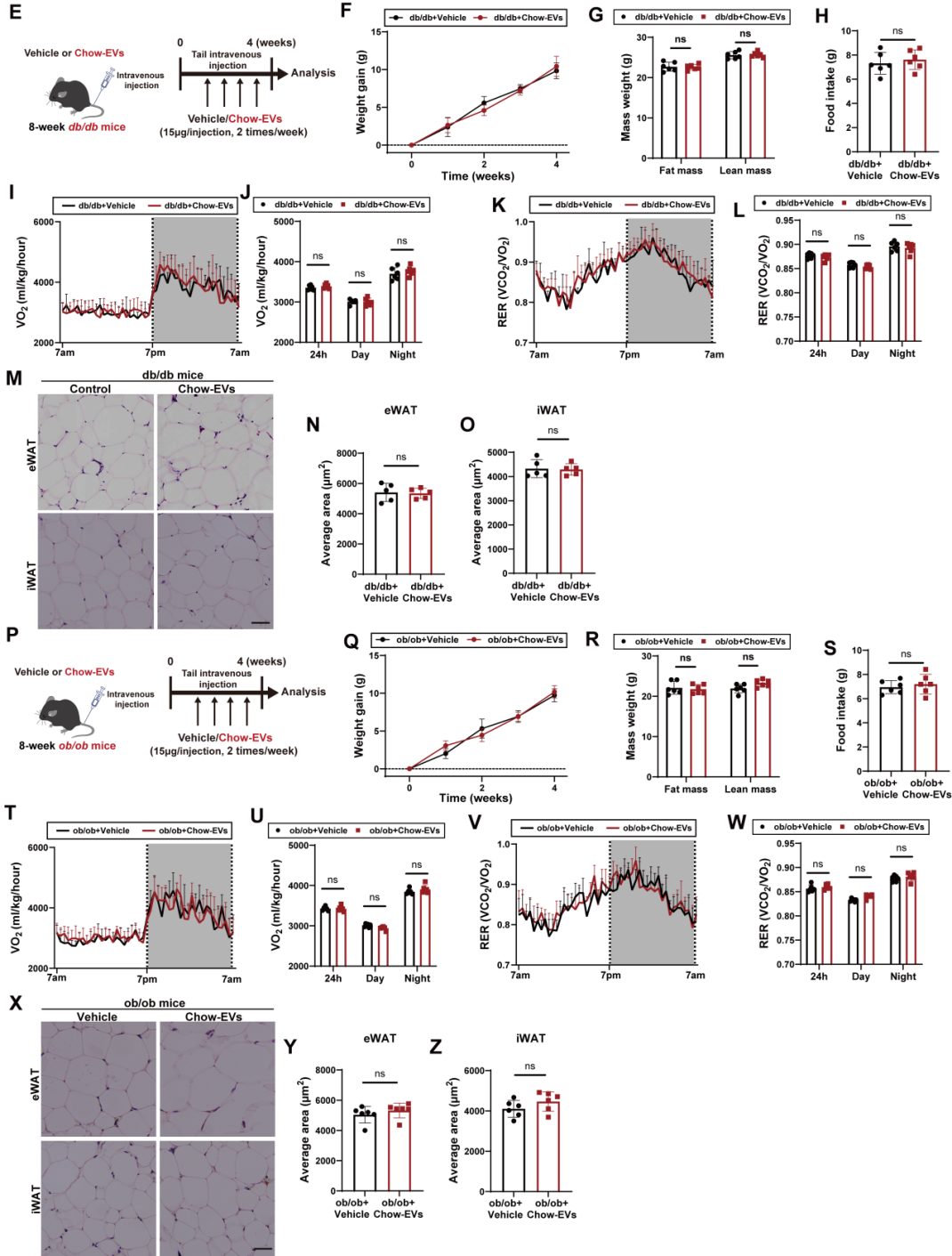
为进一步明确AT-EVs的作用是否依赖瘦素信号,研究团队向瘦素基因缺陷型肥胖小鼠(ob/ob小鼠,无法分泌瘦素)与瘦素受体缺陷型肥胖小鼠(db/db小鼠,无法接收瘦素信号)静脉注射chow-EVs或vehicle,结果显示:与对照组相比,这两种注射chow-EVs小鼠的体重、脂肪量、摄食量、能量消耗等指标均无明显变化。该结果表明,AT-EVs对能量平衡的调控作用依赖于瘦素信号通路。
图2. chow-EVs在ob/ob或db/db小鼠中均未引起代谢改善
2.AT-EVs对下丘脑瘦素-黑皮质素信号通路的调控至关重要
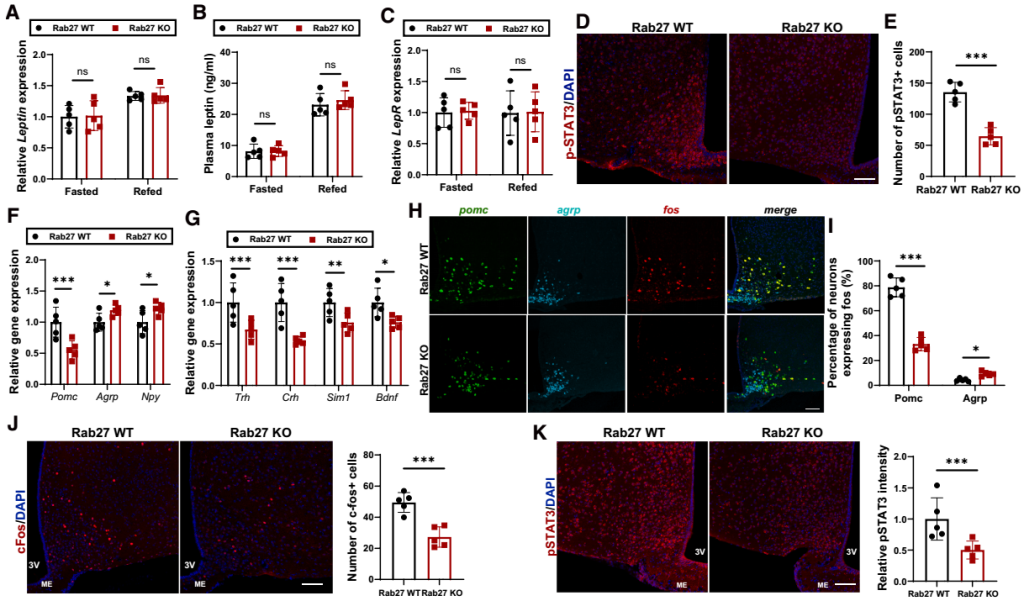
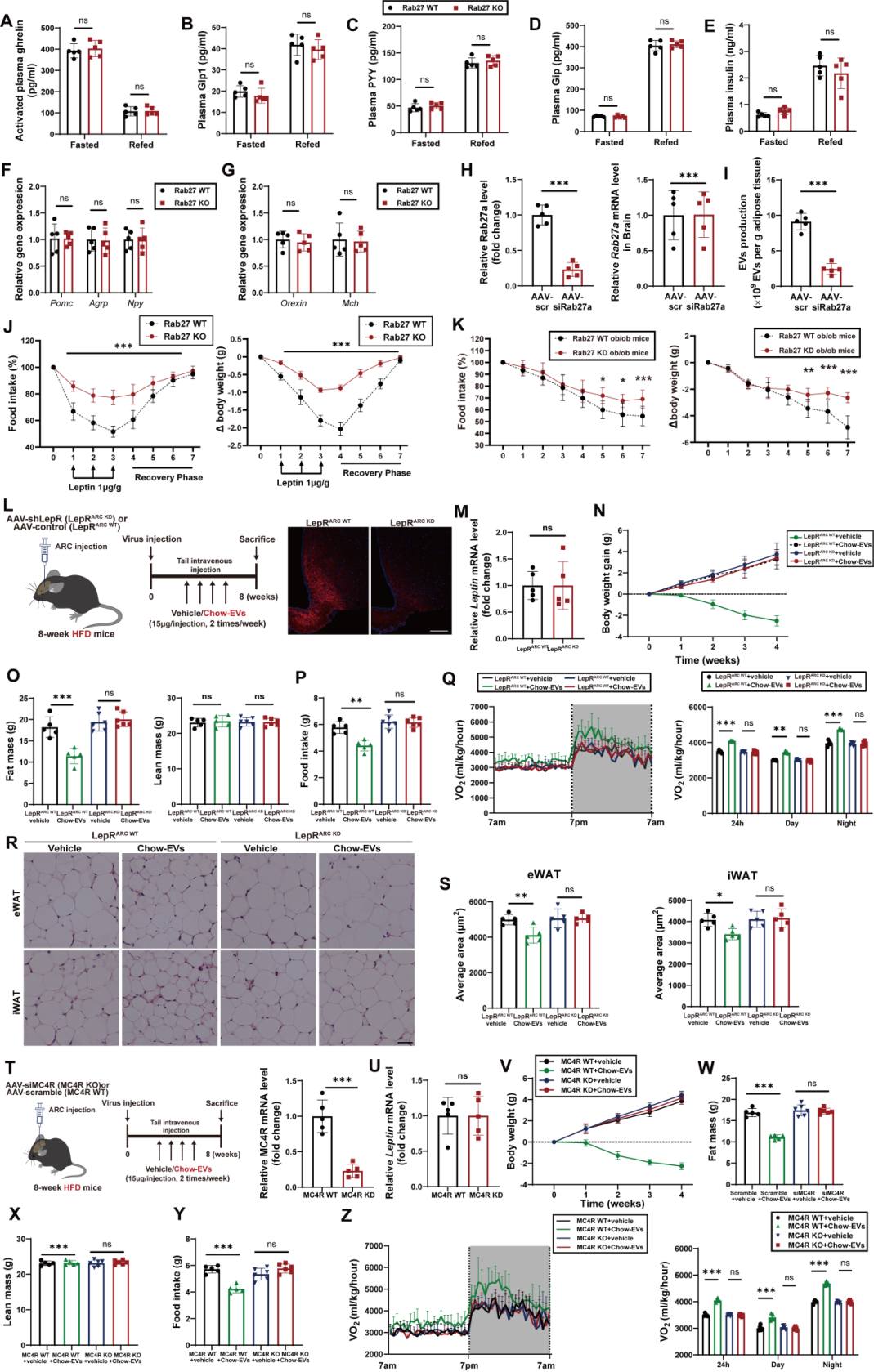
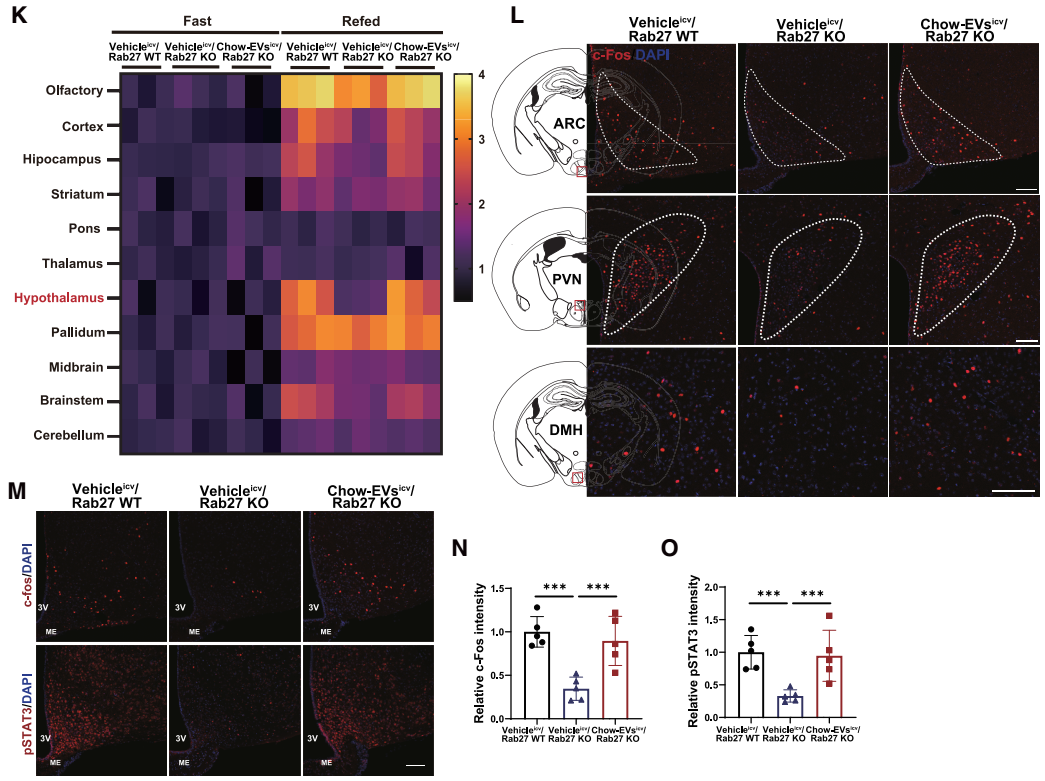
研究人员进一步探究了AT-EVs在瘦素相关效应机制中的作用。团队首先排除了“AT-EVs影响瘦素生成或外周激素水平”的可能性:在禁食与复食条件下,Rab27 KO小鼠与Rab27 WT小鼠的WAT瘦素基因表达和血浆瘦素水平无差异;同时,血浆中其他影响摄食和能量消耗的外周激素水平也无明显变化。由于瘦素对能量平衡的调控作用主要通过下丘脑LepR神经元介导,团队进一步检测下丘脑的瘦素信号发现:在禁食和复食条件下,Rab27 KO小鼠与Rab27 WT小鼠的下丘脑LepR基因表达无差异,但复食后Rab27 KO小鼠下丘脑内瘦素作用生物标志物——磷酸化信号转导与转录激活因子3(pSTAT3)的蛋白水平显著降低,提示瘦素信号传导受阻。
由于瘦素信号通过激活下丘脑黑皮质素系统抑制食欲、促进能量消耗,团队进一步检测了该通路关键神经肽的表达:复食条件下,Rab27 KO小鼠的促食欲神经肽Agrp(刺鼠相关蛋白)和Npy(神经肽Y)mRNA水平显著升高,抑食欲神经肽Pomc(阿片促黑素原)mRNA水平显著降低。同时,黑皮质素4受体(MC4R)通路下游的效应因子的表达也明显下降;而禁食条件下,两组小鼠的神经肽表达无差异,提示AT-EVs主要在“能量充足”状态下调控神经肽平衡。此外,团队通过多重原位荧光杂交(MERFISH)观察神经元活性发现,复食后,与Rab27 WT小鼠相比,Rab27 KO小鼠的激活态POMC神经元(fos+ Pomc)与激活态MC4R神经元(fos+ MC4R)数量减少,激活态AgRP神经元(fos+ Agrp)数量增加。综上,AT-EVs分泌受阻会抑制下丘脑瘦素-黑皮质素信号的激活。
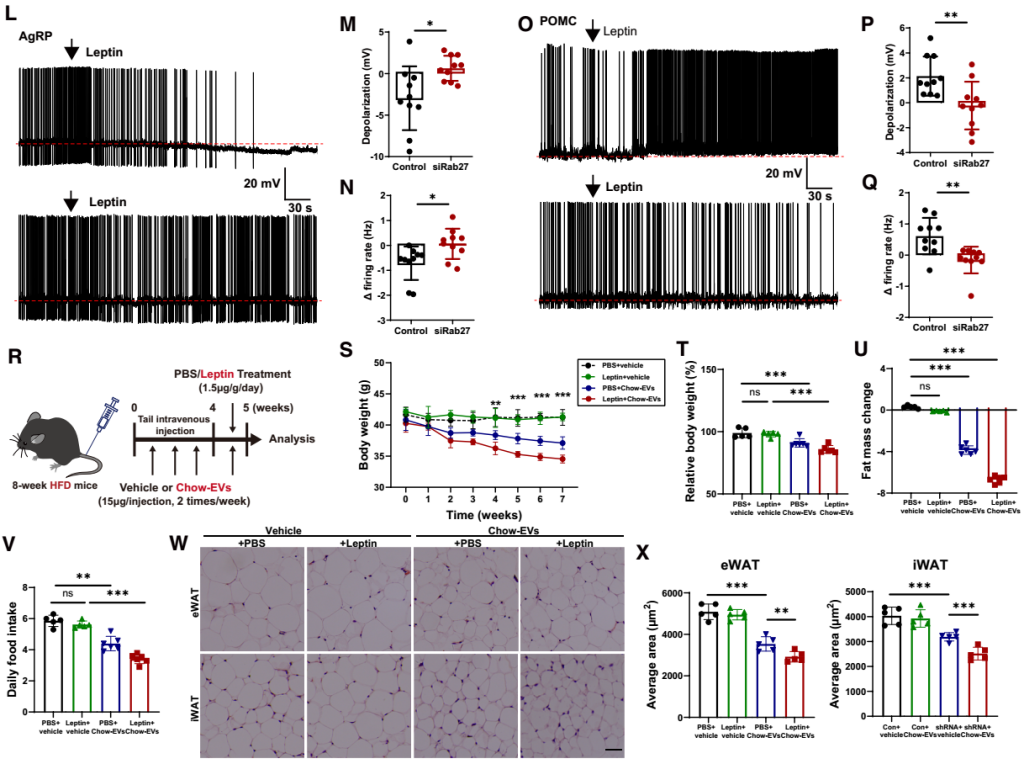
为明确AT-EVs对神经元“瘦素应答能力”的影响,研究团队进行了系列功能实验:首先,向chow喂养的Rab27 KO与Rab27 WT小鼠单次腹腔注射瘦素(3 mg/kg),结果显示KO小鼠弓状核(ARC,瘦素-黑皮质素通路上游起始脑区)的c-Fos(神经元激活标志物)与pSTAT3荧光强度显著低于WT小鼠(提示中枢对瘦素的应答减弱)。由于瘦素通过抑制AgRP神经元、激活POMC神经元来激活下丘脑黑皮质素系统,团队进一步探究了AT-EVs在这些应答中的作用,通过神经元特异性荧光报告小鼠结合电生理记录发现:向小鼠脂肪组织注射表达Rab27a小干扰RNA(siRNA)的AAV(抑制AT-EVs分泌)后,小鼠的瘦素诱导超极化显著减弱,AgRP神经元放电频率受抑;POMC神经元的“瘦素诱导放电频率增加”效应也明显降低——表明AT-EVs分泌受阻直接损害了AgRP与POMC神经元对瘦素的应答。
为进一步验证AT-EVs对瘦素功能的必要性,团队进行了外源性瘦素干预实验:向Rab27 WT小鼠注射瘦素可显著降低其摄食量与体重,但这一效应在Rab27 KO小鼠中显著减弱;在保留中枢瘦素敏感性的ob/ob小鼠中,先通过AAV敲低脂肪组织的Rab27a(Rab27a KD ob/ob小鼠),2周后连续7天腹腔注射瘦素,其“抑制摄食、减重”的效应相比Rab27a WT ob/ob小鼠显著衰减。更重要的是,向DIO小鼠补充chow-EVs后,再注射瘦素,可观察到“体重降低、脂肪量减少、脂肪细胞体积减小、摄食量下降”的效应——而未补充chow-EVs的DIO小鼠对瘦素无应答,证实AT-EVs可恢复肥胖小鼠的瘦素应答。
为明确“下丘脑瘦素-黑皮质素通路”是AT-EVs发挥作用的必需通路,团队利用AAV敲低不同脑区的关键分子:向DIO小鼠的ARC注射AAV-shLepR(LepRARC KD),或向室旁核(PVN,瘦素-黑皮质素通路下游效应脑区)注射AAV-shMC4R(MC4R KD)。结果显示,在LepRARC KD或MC4R KD小鼠中,chow-EVs的“体重、脂肪量和摄食量降低,能量消耗增加”效应完全消失;而在对照组小鼠(未敲低相应分子)中,chow-EVs仍能有效改善代谢表型,表明抑制瘦素-下丘脑黑皮质素轴会减弱AT-EVs介导的能量平衡调控。综上,AT-EVs对维持下丘脑黑皮质素系统的瘦素应答至关重要。
图3. AT-EVs对调节下丘脑瘦素-黑皮质素信号通路至关重要
图4. AT-EVs通过调节下丘脑黑皮质素系统对瘦素的应答来调控能量稳态
3.AT-EVs通过直接作用于下丘脑瘦素-黑皮质素回路调控能量稳态
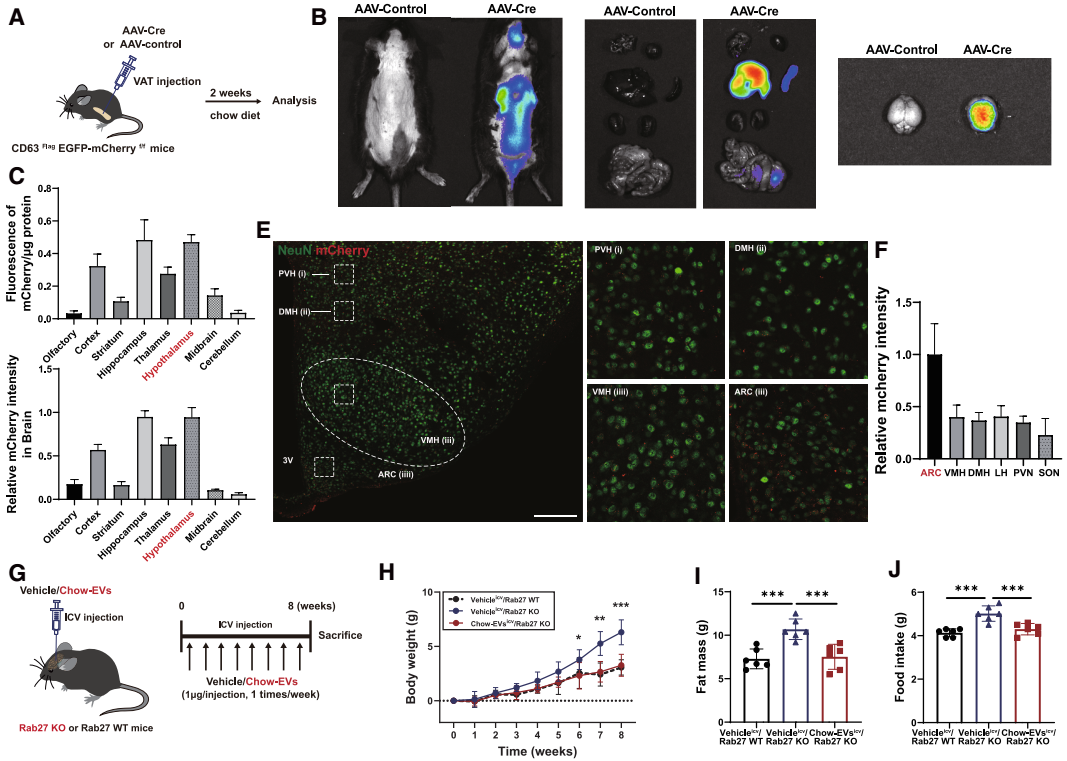
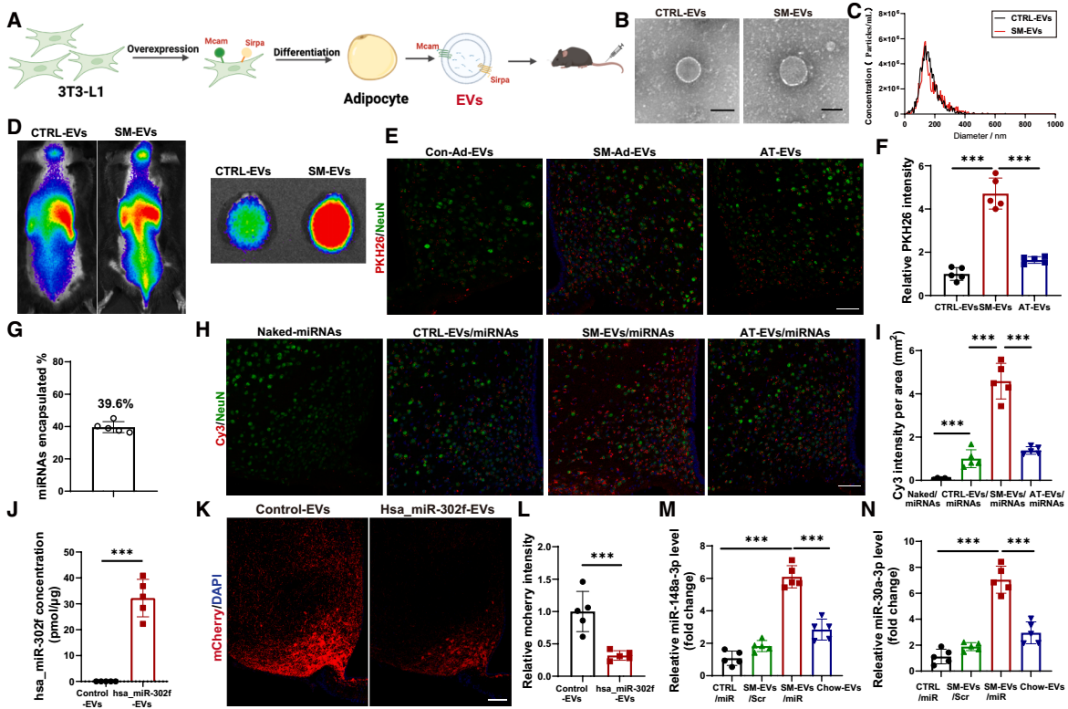
研究团队进一步通过脂肪组织特异性EV示踪技术方法,明确了AT-EVs的中枢靶向特性。首先,团队向CD63Flag EGFP-mCherryf/f报告小鼠的脂肪组织注射表达Cre重组酶的AAV,Cre可切割小鼠报告基因的loxP位点,解除对mCherry红色荧光蛋白基因表达的抑制,使EV膜蛋白CD63携带mCherry荧光标签,从而标记AT-EVs。AAV注射4周后,在小鼠大脑中检测到显著的mCherry荧光信号,提示AT-EVs可转运至CNS。为进一步定位AT-EVs在脑内的分布,团队向WT小鼠脂肪组织注射表达棕榈酰化mCherry(palm-mCherry,可使mCherry锚定在EV膜上)的AAV,2周后全脑荧光示踪和筛选显示:mCherry信号在海马体、皮质等脑区均有分布,但在下丘脑的富集程度最高,包括下丘脑的ARC、PVN和背内侧下丘脑(DMH)等核团,其中ARC的信号强度最高。此外,通过免疫荧光共定位实验进一步发现,mCherry与神经元标志物NeuN的共定位程度显著高于小胶质细胞标志物IBA-1或星形胶质细胞标志物GFAP,表明AT-EVs对神经元具有显著的偏好性;且在ARC的LepR+神经元中可检测到mCherry信号,印证了AT-EVs直接作用于下丘脑瘦素-黑皮质素信号通路这一观点。
为明确AT-EVs的“中枢作用”是否直接介导能量稳态调控,研究团队进行了脑室内(i.c.v.)补充AT-EVs实验:向Rab27 KO小鼠i.c.v.注射chow-EVs,(chow-EVi.c.v.-Rab27 KO),结果显示,与vehicle处理的Rab27 KO小鼠相比,chow-EVi.c.v.-Rab27 KO小鼠体重显著降低、日摄食量减少、能量消耗增加、RER下降、脂肪细胞体积缩小。随后通过全脑c-Fos筛选进一步证实,复食后Rab27 KO小鼠多个脑区(包括嗅球、海马体、下丘脑等)c-Fos信号显著降低(神经元活性抑制),而i.c.v.补充chow-EVs可完全恢复这些脑区(尤其ARC、DMH、PVN等下丘脑核团)的神经元活性,表明中枢补充AT-EVs可缓解AT-EVs抑制引起的神经元活性异常。此外,chow-EVi.c.v.-Rab27 KO小鼠的下丘脑瘦素-黑皮质素通路相关基因表达与Rab27 WT小鼠相当,表明中枢补充AT-EVs可恢复因AT-EVs缺失导致的中枢瘦素信号受损。上述结果充分证实,AT-EVs可直接作用于下丘脑瘦素-黑皮质素回路来调控能量稳态。
图5. AT-EVs直接作用于CNS来调节能量稳态
4.miRNAs是AT-EVs调控能量稳态的关键介质
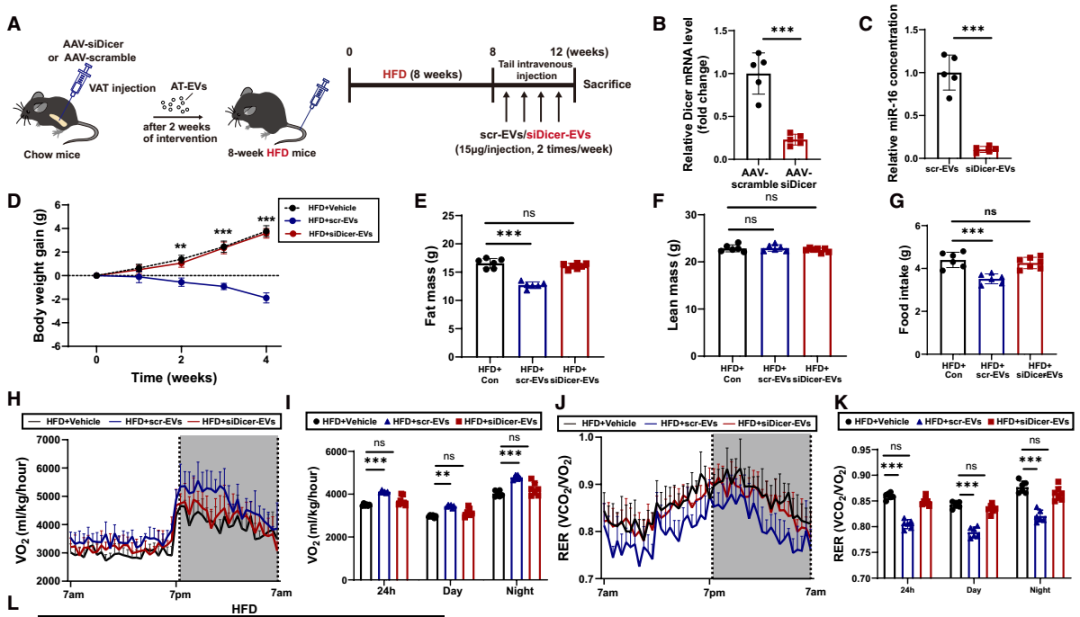
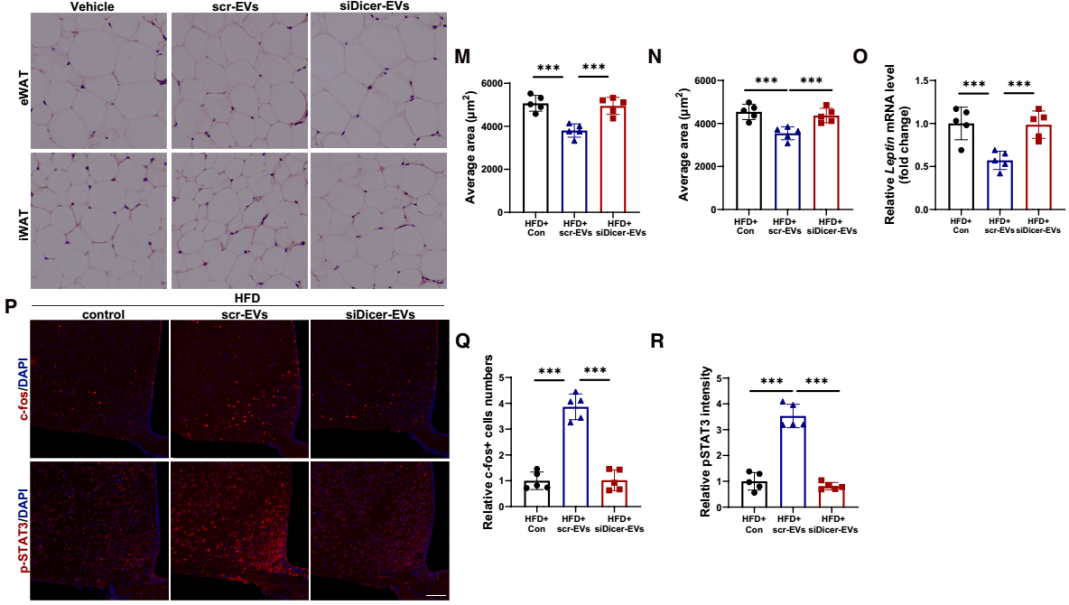
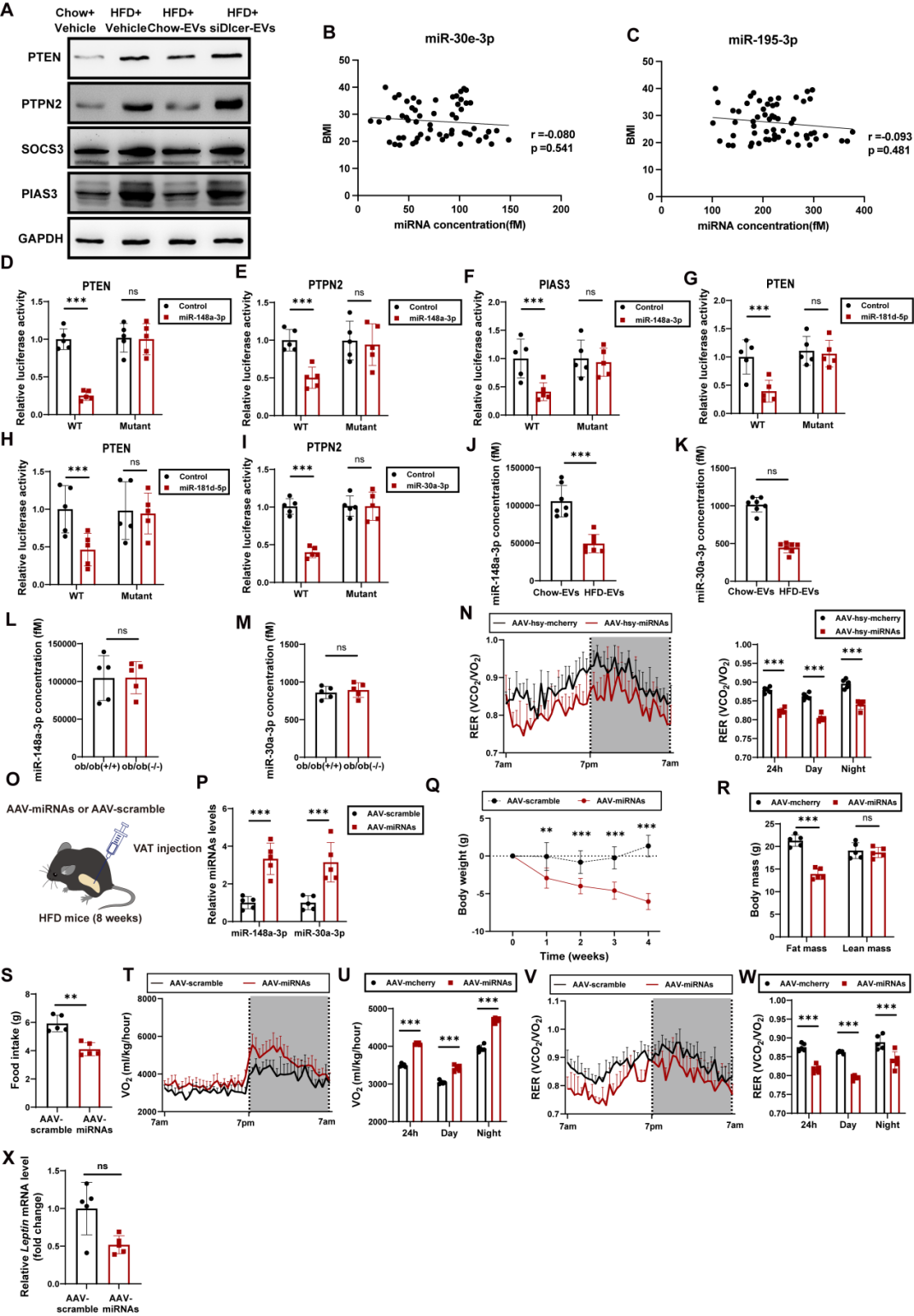
EVs可携带蛋白质、核酸等多种货物分子,其中miRNA是主要成分。为探究miRNA是否介导AT-EVs的能量调控作用,研究团队构建了脂肪组织miRNA生成缺陷模型小鼠:向chow喂养小鼠的脂肪组织注射表达Dicer siRNA的AAV——Dicer是miRNA加工成熟的关键酶,其缺失可导致EVs内miRNA含量显著降低;对照组则注射AAV-scramble。随后,从上述小鼠脂肪组织中提取EVs(分别命名为siDicer-EVs与scr-EVs),静脉注射到DIO小鼠中。实验结果显示:与vehicle处理的DIO小鼠相比,scr-EVs可显著降低DIO小鼠体重与脂肪量、减少日摄食量、增加能量消耗、降低RER,同时减小脂肪细胞体积并降低瘦素mRNA水平;而siDicer-EVs处理对上述指标无明显影响。提示AT-EVs的代谢调控效应依赖于其携带的miRNA。进一步的瘦素应答实验显示,scr-EVs处理的DIO小鼠在腹腔注射瘦素后,ARC中的c-Fos强度与pSTAT3表达显著升高(瘦素敏感性恢复),而siDicer-EVs和vehicle处理的DIO小鼠无此效应。以上结果表明,miRNA介导了AT-EVs的瘦素增敏和抗肥胖作用。
图6. miRNAs介导AT-EVs调控能量稳态的作用
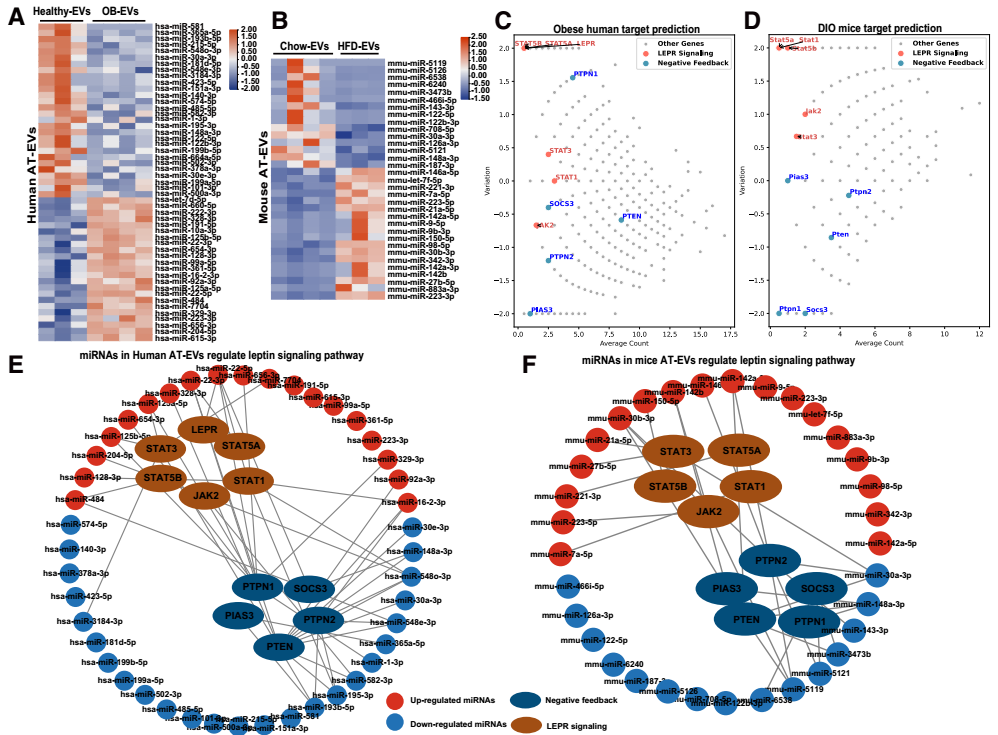
为鉴定AT-EVs中瘦素增敏性miRNA,研究团队通过深度RNA测序分析了人和小鼠“健康EVs”与“肥胖EVs”的miRNA表达谱,随后通过靶基因预测与功能富集分析发现,肥胖AT-EVs中下调的miRNA通过靶向瘦素受体信号通路的负反馈调控因子(如SOCS3、PIAS3、PTEN、PTPN2)来维持瘦素敏感性;而肥胖AT-EVs中上调的miRNA则通过靶向瘦素受体下游效应因子(如LepR、JAK2、STAT3)来抑制瘦素信号。
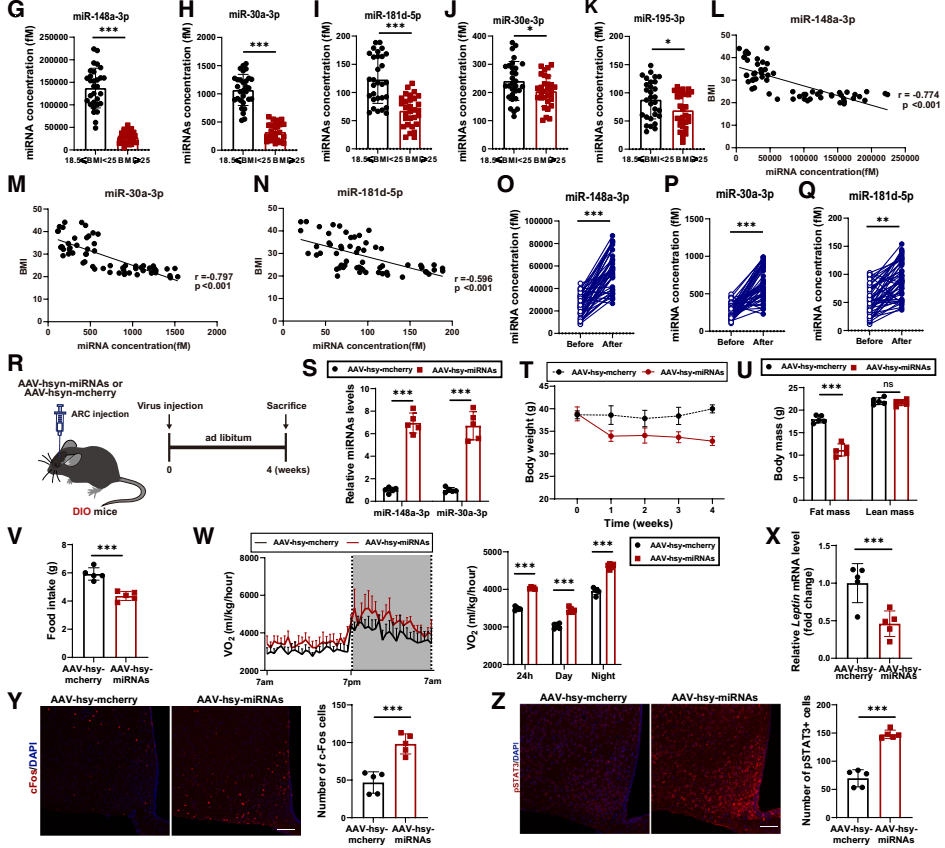
为建立miRNA与人类肥胖的关联性,研究团队进一步开展了两项临床队列研究:队列1包含30名超重/肥胖者与30名年龄、性别匹配的正常体重健康者,结果显示肥胖者血清EVs中的5种瘦素增敏性miRNA水平显著低于正常者,其中miR-148a-3p、miR-30a-3p和miR-181d-5p的水平与BMI(体质指数)呈负相关;队列2包含50名接受Roux-en-Y胃旁路手术(减重代谢手术)的肥胖患者,术后6个月,患者血清EVs中上述三种miRNA的水平显著升高。双荧光素酶报告基因实验进一步证实,这些miRNA可直接靶向SOCS3、PIAS3、PTEN;小鼠实验也显示,DIO小鼠血清EVs中miR-148a-3p和miR-30a-3p水平显著降低,而ob/ob小鼠的这些miRNA维持在较高水平。进一步验证了这些miRNA在AT-EVs中的瘦素增敏作用。
为明确其治疗潜力,研究团队向DIO小鼠的ARC或脂肪组织中注射过表达miR-148a-3p和miR-30a-3p的AAV,结果显示,无论在中枢还是外周,过表达这两种miRNA,均可显著降低DIO小鼠的体重、摄食量、RER、瘦素mRNA水平,并增加其能量消耗。证实miR-148a-3p和miR-30a-3p可增强瘦素敏感性并抑制肥胖。
图7. 健康EVs中富集的miRNA表达谱发挥瘦素增敏作用
5.利用工程化AT-EVs靶向CNS递送瘦素增敏性miRNA可改善肥胖
接下来,研究团队探索了上述发现在治疗肥胖方面的转化潜力。首先,团队明确了AT-EVs靶向CNS的分子机制。对比不同脂肪组织来源的EVs,发现小鼠的内脏脂肪组织(VAT)来源的EVs具有更优的下丘脑靶向性,并进一步确定是VAT中的脂肪细胞[而非“非脂肪细胞”的基质血管组分细胞(SVCs)]来源的EVs(Ad-EVs)能够转运至下丘脑。接着,研究团队鉴定了介导AT-EVs中枢靶向的关键膜蛋白。对比AT-EVs与肝脏来源EVs(L-EVs)的靶向性发现,AT-EVs在下丘脑的荧光信号显著强于L-EVs;若用胰蛋白酶预处理AT-EVs(切割膜蛋白),其下丘脑靶向能力显著下降(证实膜蛋白是靶向关键)。进一步通过无偏倚鸟枪法质谱法对AT-EVs和L-EVs进行蛋白质组学分析,筛选出115种在AT-EVs中独特或高度富集的蛋白,其中71种为膜蛋白。随后,团队通过“功能注释”与“siRNA敲低筛选”,最终确定两种关键膜蛋白——SIRPα(信号调节蛋白α)与MCAM(黑色素瘤细胞粘附分子):向小鼠脂肪组织注射表达SIRPα或MCAM siRNA的AAV后,其来源EVs的下丘脑靶向能力显著降低——证实SIRPα和MCAM是AT-EVs靶向CNS的分子基础。
基于上述发现,研究团队构建了中枢靶向性工程化EVs(SM-EVs):用携带编码SIRPα和MCAM质粒的慢病毒(LV-SM)或其对照(LV-control)转染3T3-L1脂肪前体细胞,并纯化来自分化3T3-L1脂肪细胞的EVs(分别为SM-EVs和control-EVs)。通过TEM与纳米颗粒追踪分析显示,SM-EVs与control-EVs的物理性质无差异,但SM-EVs的SIRPα与MCAM蛋白水平显著升高。为验证SM-EVs的中枢靶向性,研究团队向小鼠静脉注射PKH26(亲脂性红色荧光染料)标记的SM-EVs、control-EVs与天然AT-EVs,体外荧光成像显示,SM-EVs在CNS的荧光信号显著强于control-EVs与天然AT-EVs,且荧光信号主要富集在下丘脑。进一步通过Cy3(水溶性红色荧光染料)标记miRNA的递送实验证实,SM-EVs可将Cy3标记的人特异性miRNA(hsa_miR-302f,无小鼠同源物)高效递送至下丘脑,其效率显著高于control-EVs与裸miRNA(无EV包裹)。为了验证由SM-EVs递送到下丘脑的miRNAs是否能执行其调节功能,团队向小鼠ARC注射携带hsa_miR-302f的3'UTR mCherry报告基因的AAV,3天后再注射装载hsa_miR-302f的SM-EVs,2周后检测结果显示,小鼠ARC的hsa_miR-302f水平升高,mCherry荧光强度降低——证实SM-EVs可将miRNA递送至下丘脑并发挥基因调控功能。
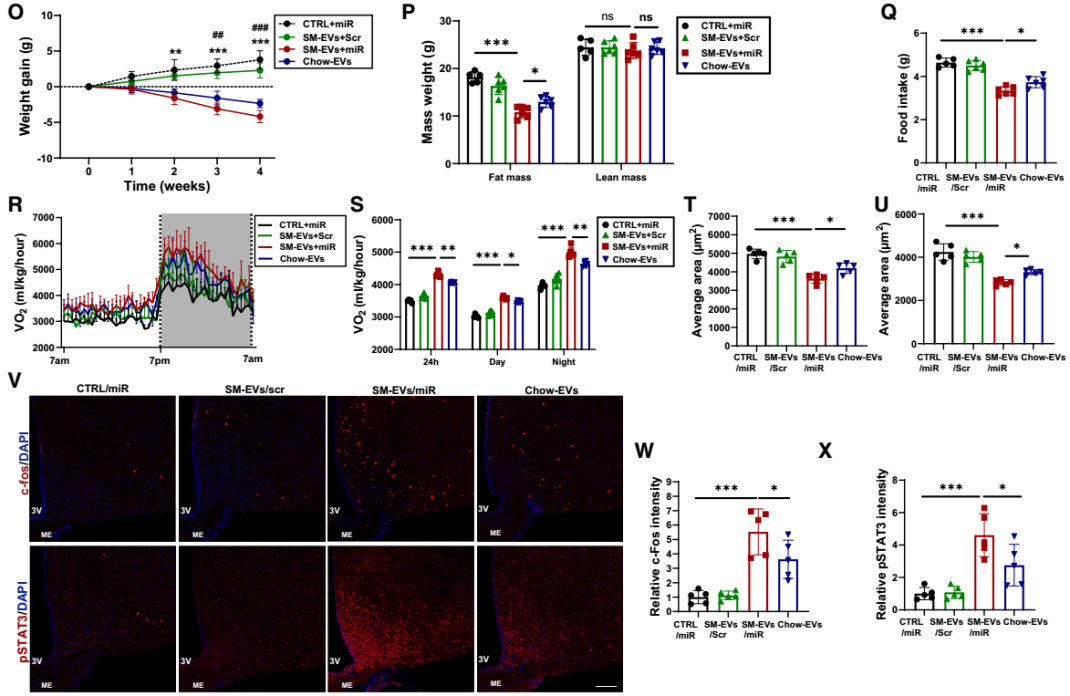
最后,研究团队评估了SM-EVs向CNS递送瘦素增敏性miRNA的抗肥胖效应。向DIO小鼠静脉注射裸miR-148a-3p/miR-30a-3p或SM-EVs包裹的miR-148a-3p/miR-30a-3p。处理4周后,与裸miRNA组相比,SM-EVs/miRNA组小鼠的体重显著降低、日摄食量减少、能量消耗增加,且中枢瘦素敏感性增强。这些结果表明,靶向CNS递送瘦素增敏性miRNA是增强瘦素敏感性和治疗肥胖的有效策略。
图8. 利用工程化EVs靶向CNS递送瘦素增敏性miRNAs可缓解肥胖
总结
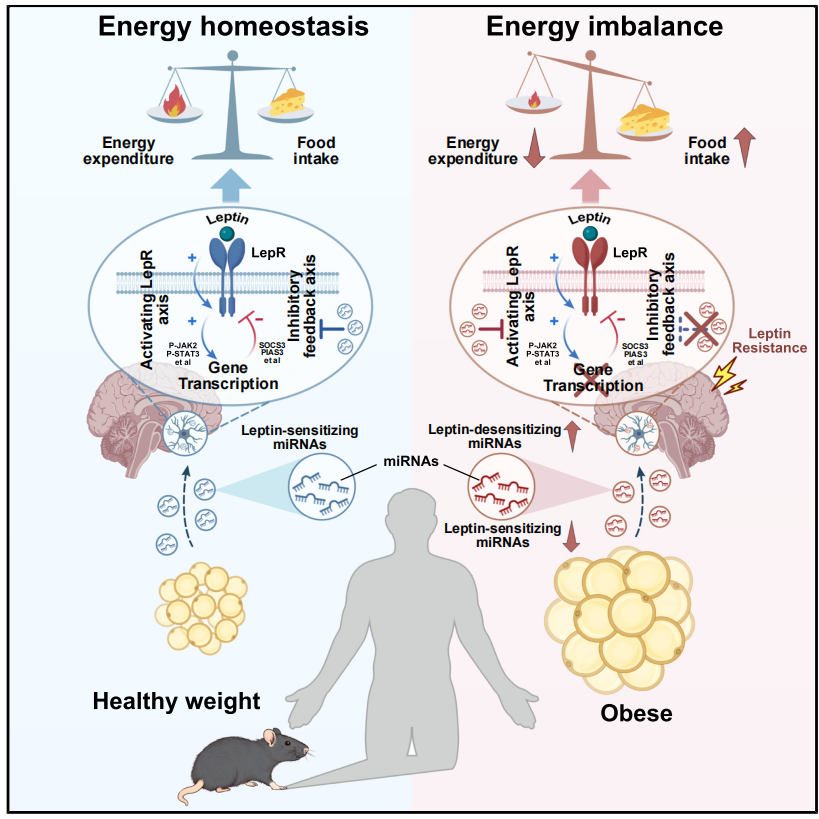
该研究通过将研究范式从“大脑中心视角”转向“器官间通讯框架”,极大地推进了对中枢瘦素抵抗机制的理解。研究首次证明,脂肪组织与大脑之间正常通讯的破坏是构成中枢瘦素抵抗的主要机制。在此背景下,研究团队揭示了AT-EVs及其携带的miRNA可作为调节下丘脑回路对瘦素应答的新型调控因子。其中,Ad-EVs内一组miRNA被鉴定为瘦素增敏剂,它们的缺失是驱动中枢瘦素抵抗的关键因素。重要的是,团队构建了经特定Ad-EVs膜蛋白修饰的工程化EVs,可将瘦素增敏性miRNA靶向递送至CNS,增强肥胖小鼠的中枢瘦素敏感性并诱导其显著减重。这些发现不仅拓宽了我们对脂肪-脑轴的理解,更为提升中枢瘦素敏感性、治疗肥胖提供了具有前景的治疗策略。
总结图
南京大学医学院附属鼓楼医院王进副研究员为该论文的唯一第一作者。南京大学医学院附属鼓楼医院毕艳主任医师和南京大学李靓教授为该论文的共同通讯作者。该成果得到了国家自然科学基金原创探索项目和江苏省自然科学基金等项目的资助。
毕艳/主任医师
毕艳,南京大学医学院附属鼓楼医院副院长,临床医学研究院院长,南京大学代谢病研究所所长,内分泌科学科带头人,主任医师,教授,博士生导师。国家“高层次人才支持计划”科技创新领军人才。中华医学会糖尿病学分会常务委员,中华医学会糖尿病学分会肥胖与糖尿病学组副组长,江苏省医学会糖尿病学分会候任主任委员,江苏省老年医学学会老年内分泌分会主任委员,中华糖尿病杂志编委。聚焦糖尿病和肥胖诊疗新技术的基础临床及转化研究,以第一及通讯作者在Cell Metabolism、Diabetes Care、Nature Communications、Journal of Hepatology、Diabetes等杂志发表SCI论著101篇,总影响因子550分。主持国家科技重大专项、国家自然科学基金重点项目、原创探索项目等。获批国际/中国发明专利/软著授权10项,并实现成果转化。相关研究获江苏医学科技奖一等奖,中华医学会糖尿病学分会杰出青年科学研究奖。主编专著《内分泌代谢病临床思维训练》。
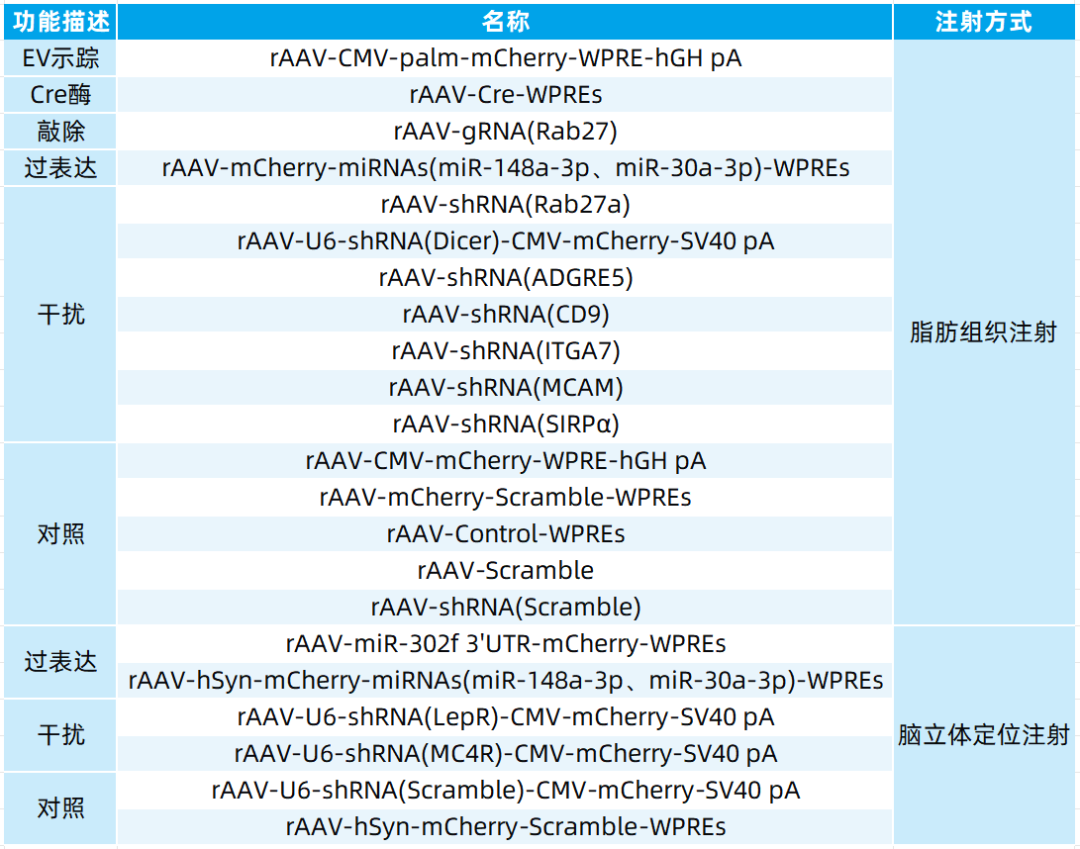
本文使用的病毒产品均来自枢密科技,列表如下:

了解产品及服务
请扫码添加客服微信:BrainVTA2020

市场:027-65023363 行政/人事:027-62439686 邮箱:marketing@brainvta.com 客服:18140661572(活动咨询、售后反馈等)
销售总监:张经理 18995532642 华东区:陈经理 18013970337 华南区:王经理 13100653525 华中/西区:杨经理 18186518905 华北区:张经理 18893721749
地址:中国武汉东湖高新区光谷七路128号中科开物产业园1号楼
Copyright © 武汉枢密脑科学技术有限公司. All RIGHTS RESERVED.
鄂ICP备2021009124号 DIGITAL BY VTHINK