2023-03-17 阅读量:30654
阿尔茨海默病(AD)是最常见的与年龄相关的神经退行性疾病。AD的主要临床表现是进行性认知功能下降。病理学上,AD的特征是突触功能障碍、淀粉样斑块和神经原纤维缠结的聚集。突触功能障碍是AD症状的主要原因。此外,突触功能障碍是AD非常早期的特征,且在出现认知症状数年之前发生。
突触素Ⅰ是定位于突触囊泡(SVs)细胞质表面的神经元特异性磷酸蛋白。它是突触前末梢SVs动力学的关键调节器。在静息状态下,突触素Ⅰ通过与磷脂和F-actin细胞骨架相互作用,在储备池中聚集SVs。刺激时,突触素Ⅰ在Ser9处磷酸化并与SVs分离。刺激后,突触素Ⅰ去磷酸化,并重新聚集SVs。因此,突触前终末SVs的再循环需要突触素Ⅰ的正常功能。在AD患者中,大脑中突触素Ⅰ的表达降低,并伴有突触功能障碍,提示突触素Ⅰ的功能障碍可能促使AD的突触功能障碍。然而,AD中突触功能障碍的分子机制仍不清楚。
哺乳动物天冬酰胺内肽酶(AEP)是一种溶酶体半胱氨酸蛋白酶,可切割天冬酰胺残基C端侧蛋白质底物。AEP在酸性条件下连续去除C-和N-末端的前体肽而被激活。武汉大学人民医院神经内科张振涛教授课题组前期研究结果表明AEP在大脑中以年龄依赖的方式被激活,并切割淀粉样前体蛋白(APP)和tau,促进AD发病过程中β-淀粉样蛋白(Aβ)和tau的沉积。2022年4月20日,张振涛教授团队在Aging Cell发表题为“A synapsin Ⅰ cleavage fragment contributes to synaptic dysfunction in Alzheimer's disease”的研究论文,本文中,作者发现突触素Ⅰ作为最重要的突触蛋白被 AEP 切割。研究人员证明AEP在体外和体内均能切割突触素Ⅰ的N82处,产生突触素Ⅰ(83-705)片段(命名为C83片段)。该片段在神经元中异常分布,在体外和体内均诱导突触功能障碍。野生型小鼠海马中过表达AEP导致突触素Ⅰ C83片段的产生,并诱导突触功能障碍和认知缺陷。此外,突触素Ⅰ C83片段在tau P301S转基因小鼠或野生型小鼠海马中的过表达均促进突触功能障碍和认知损伤。总的来说,本文研究结果表明AEP介导的突触素Ⅰ片段化导致AD的突触功能障碍。这些发现提示了AD中突触功能障碍的新机制。

1.突触素Ⅰ是AEP的底物
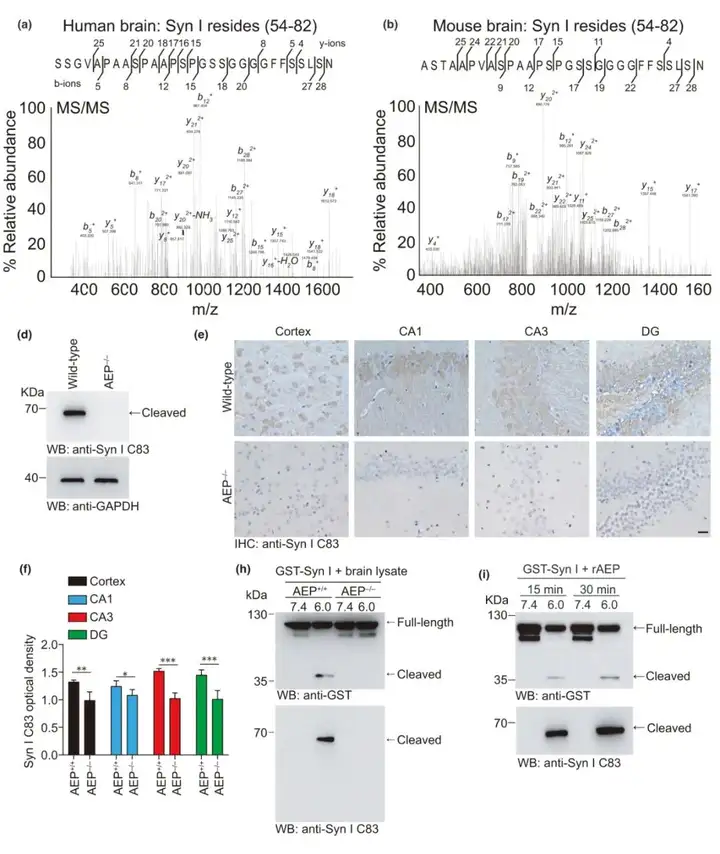
为研究突触功能障碍的分子机制,研究人员通过质谱分析AD患者的脑组织,结果发现AD患者大脑中存在突触素Ⅰ在N82位点结尾的片段(即Synapsin Ⅰ C83片段)。由于AEP是唯.一特异性切割天冬酰胺残基C端侧的蛋白酶,于是作者推测该片段是由AEP介导的切割产生的。为验证突触素Ⅰ能被AEP切割,质谱分析野生型和AEP敲除(AEP–/–)小鼠脑组织,结果显示野生型小鼠脑组织裂解物中鉴定出突触素Ⅰ的相同片段。此外,蛋白质组学分析表示相比于野生型小鼠,AEP–/–小鼠脑提取物中该片段的信号要低得多。这些结果表明AEP可能切割脑内突触素Ⅰ。进一步探究脑内Synapsin Ⅰ C83片段的存在,作者纯化出特异性识别Synapsin Ⅰ C83片段(仅由AEP切割产生)而非全长Synapsin Ⅰ 的抗体,利用该抗体通过蛋白质免疫印迹和免疫组化方法在野生型小鼠而非AEP–/–小鼠中检测到Synapsin Ⅰ C83片段的存在。为证实AEP对突触素Ⅰ的切割,研究人员将GST标记突触素Ⅰ分别与提取自野生型和AEP–/–小鼠的脑裂解物在pH 6.0下孵育,而AEP在pH 6.0时被激活,并产生用抗GST抗体能检测到的N-末端突触素Ⅰ片段,及用抗突触素Ⅰ C83抗体检测到的C末端片段。结果提示野生型小鼠组GST标记突触素Ⅰ可被切割。进一步探讨AEP对突触素Ⅰ的切割,作者将活性AEP酶与GST标记突触素I分别孵育15分钟和30分钟。Western blot分析显示,突触素Ⅰ在AEP存在下以时间依赖的方式片段化。综上所述,这些结果表明突触素Ⅰ是AEP的底物。
2.AEP在N82处切割突触素Ⅰ
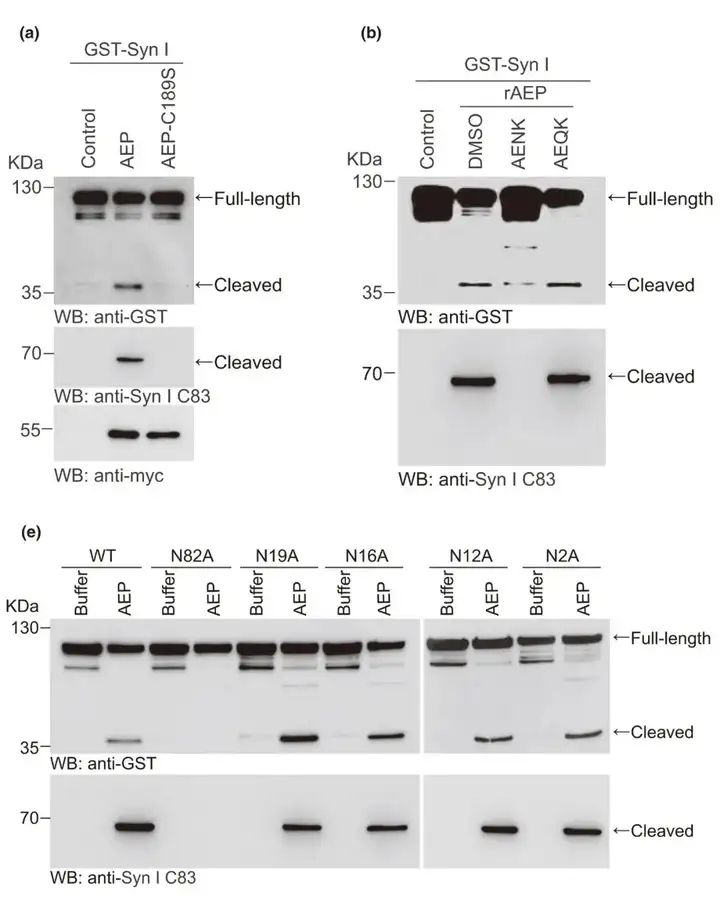
为证实AEP特异性切割突触素Ⅰ,研究人员在HEK293细胞共转染GST突触素Ⅰ和myc-AEP或myc-AEP-C189S突变体。结果显示野生型AEP强烈诱导突触素Ⅰ片段化,而AEP C189S突变体由于丧失了蛋白酶活性,不能引起对突触素Ⅰ的切割。此外,这种切割过程被AEP抑制剂AENK所阻碍。这些发现表明突触素Ⅰ C83片段是由AEP特异性产生的。前期研究结果显示来自AD患者和小鼠脑组织的质谱分析结果表明,AEP在N82处切割突触素Ⅰ。AEP选择性切割其底物C端侧的天冬酰胺残基。突触素Ⅰ的N端含有5个天冬氨酸残基(N2、N12、N16、N19和N82)。作者构建了突触素Ⅰ点突变体,将这些位点的天冬酰胺用丙氨酸取代。结果表示在N82A突变体中完全不产生突触素Ⅰ片段,这些结果表明,AEP在N82处特异性切割突触素Ⅰ,产生C83片段。
图2. AEP在N82处切割突触素Ⅰ
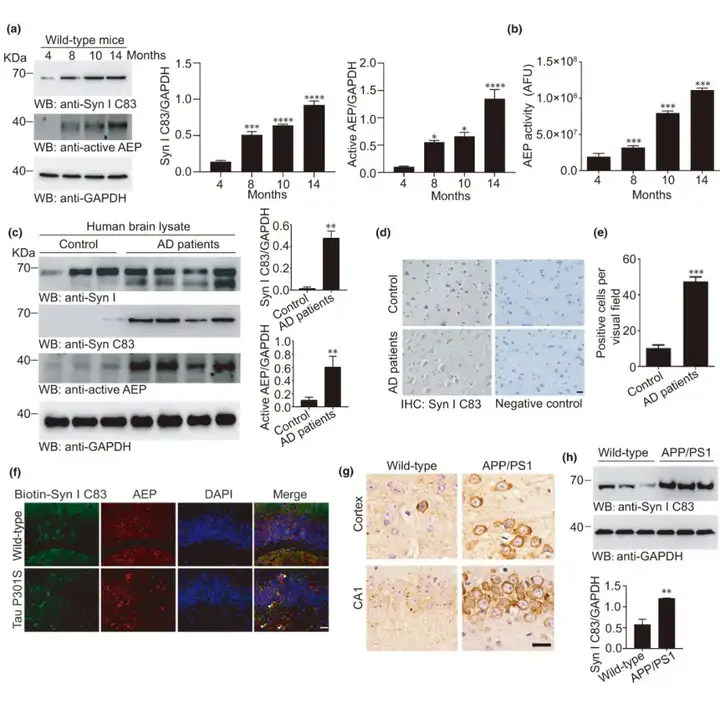
3.AEP在衰老过程中上调,并在AD脑中产生突触素Ⅰ C83片段
衰老是AD的主要风险因素。衰老过程中随着大脑内部环境逐渐酸化,AEP被激活。于是,研究人员探究了不同年龄小鼠大脑中AEP产生的突触素Ⅰ C83片段,利用抗突触素Ⅰ C83抗体检测4、8、10和14月龄小鼠脑组织裂解物中的C83片段,结果显示C83片段的量随年龄增加而增加。AEP活性测定表明,AEP在小鼠大脑中以年龄依赖的方式被激活。进一步利用蛋白质免疫印迹和免疫组化手段证实人AD脑内突触素Ⅰ C83片段的存在,结果表示突触素Ⅰ C83在人AD脑内含量丰富,但在年龄匹配的对照组脑中的水平要低得多。此外,tau P301S转基因小鼠(tau蛋白病模型,微管相关蛋白tau在细胞内形成的神经原纤维缠结是包括AD等在内的多种tau蛋白病的主要病理表现之一)海马CA1区中也检测到突触素Ⅰ C83片段的存在,但在年龄匹配的非转基因对照小鼠的脑切片中几乎没有检测到。免疫荧光染色显示AEP与突触素Ⅰ C83共定位。Western blot和免疫组化证实,来自APP/PS1转基因小鼠(成熟的AD动物模型)的脑裂解物中也富含突触素Ⅰ C83片段。因此,上述结果表明AEP在衰老过程中被激活,并切割tau P301S小鼠和APP/PS1小鼠大脑中的突触素Ⅰ。
图3. AEP被激活并切割AD脑中的突触素Ⅰ
4.体外实验证明突触素Ⅰ C83片段诱导突触功能障碍
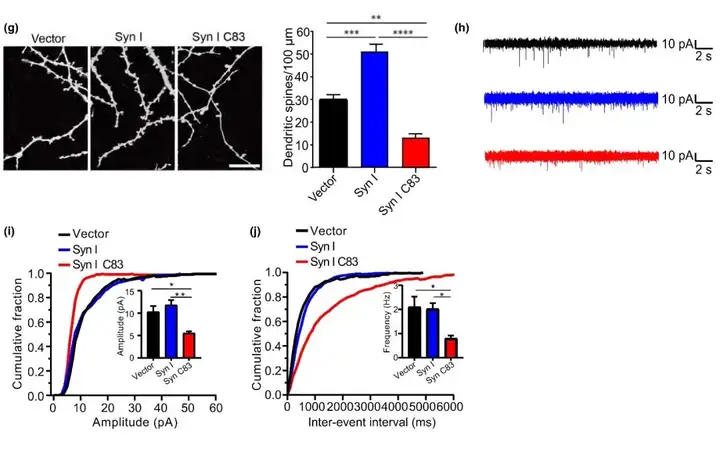
突触素Ⅰ通过与脂质和肌动蛋白细胞骨架相互作用以在储备池中聚集SVs,来调节神经递质的释放。为评估AEP介导的对突触素Ⅰ切割是否影响神经元中突触素Ⅰ的正常功能,利用分别编码EGFP、EGFP-突触素Ⅰ和EGFP-突触素Ⅰ C83的腺相关病毒(AAV)感染海马原代神经元,结果显示突触素Ⅰ呈点状分布,而突触素Ⅰ C83分布更为广泛。免疫荧光染色显示全长突触素Ⅰ与突触囊泡蛋白(定位于SVs的膜蛋白)共定位,而突触素Ⅰ C83却没有。这些结果表明C83片段不像全长蛋白那样与SVs结合。接下来,通过FM4-64(神经末梢荧光染料)标记实验探究是否突触素Ⅰ C83片段损害神经元中SVs的再循环。与过表达全长突触素Ⅰ组相比,表达突触素Ⅰ C83组的FM4-64标记减少了45%,在突触素Ⅰ C83存在下,FM4-64的释放也受损。此外,DiI染色显示,与表达全长突触素Ⅰ神经元比较,表达突触素Ⅰ C83神经元中树突棘密度更小。表明突触素Ⅰ C83片段诱导树突棘退化。这些结果表明突触素Ⅰ C83片段分布异常,损伤SVs的再循环,并诱导树突棘变性。进一步研究人员探讨突触素Ⅰ C83片段是否引起突触前缺陷,记录生长在用胶原/多聚赖氨酸(PDL)包被培养容器中的单个培养神经元的微型兴奋性自发电流(mEACs),结果提示,相比于表达全长突触素Ⅰ神经元,表达突触素Ⅰ C83神经元中mEACs频率和振幅显著降低。总的来说,上述结果表明突触素Ⅰ C83片段干扰突触传递。
图4. 突触素I C83片段的过度表达诱导突触功能障碍
5.体内实验证明过表达AEP诱导突触功能障碍与认知缺陷
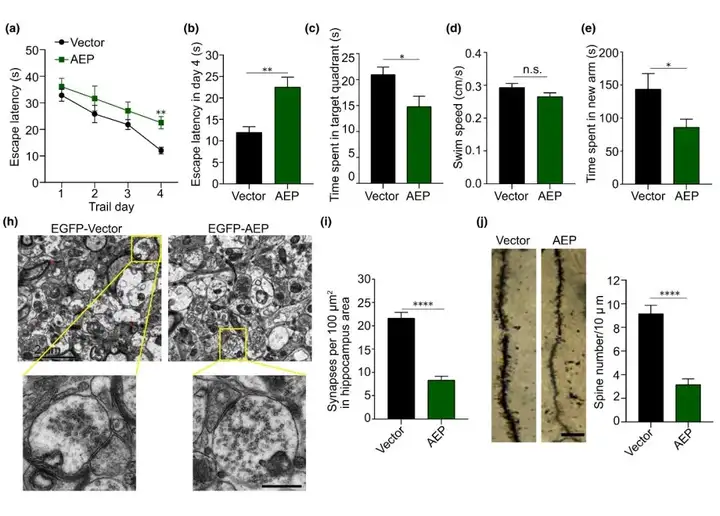
接下来作者通过体内实验评估AEP对突触和认知功能的影响。编码EGFP和EGFP-AEP的AAV分别注射到2月龄野生型小鼠海马区。三个月后,研究人员通过Morris水迷宫实验评估AEP过表达对空间学习和记忆能力的影响。结果显示在训练阶段(定位航行试验),所有小鼠找到平台的潜伏期逐渐缩短,表明有学习效果。然而,过表达AEP小鼠的学习能力严重损伤(第四天过表达AEP小鼠找到平台的时间显著长于对照组小鼠)。空间探索试验中,表达AEP的小鼠出现在目标象限的时间减少,表明记忆功能受损。所有小鼠的游泳速度相当,表明AEP的过度表达不会影响运动功能。同样,在Y迷宫实验中,表达AEP的小鼠在新异臂中花费的时间更少。这些结果表明,小鼠大脑中AEP的过度表达导致学习和记忆障碍。免疫荧光染色显示相比于对照组小鼠,过表达AEP组小鼠海马CA1和CA3区突触素Ⅰ C83信号更强。此外,EGFP-AEP与突触素Ⅰ C83片段共定位。Western blot分析显示,与对照组小鼠比较,表达AEP组小鼠海马裂解物中突触素Ⅰ C83片段的丰度增加。这些结果表明,AEP的过表达诱导体内突触素Ⅰ C83片段的产生。海马区电镜分析显示,与对照组相比,注射AAV-EGFP-AEP的小鼠海马中突触的密度降低。此外,与对照组比较,过表达AEP组SVs的分布更为分散,表明AEP的过度表达导致储备池中SVs聚集的能力降低。高尔基染色显示AEP诱导野生型小鼠树突棘的丢失。综上所述,外源AEP的过度表达导致小鼠体内突触功能障碍和认知损伤。
图5. AEP过表达诱导野生型小鼠突触素Ⅰ片段化和突触功能障碍
6.突触素Ⅰ C83片段诱导tau P301S转基因小鼠突触功能障碍和认知缺陷
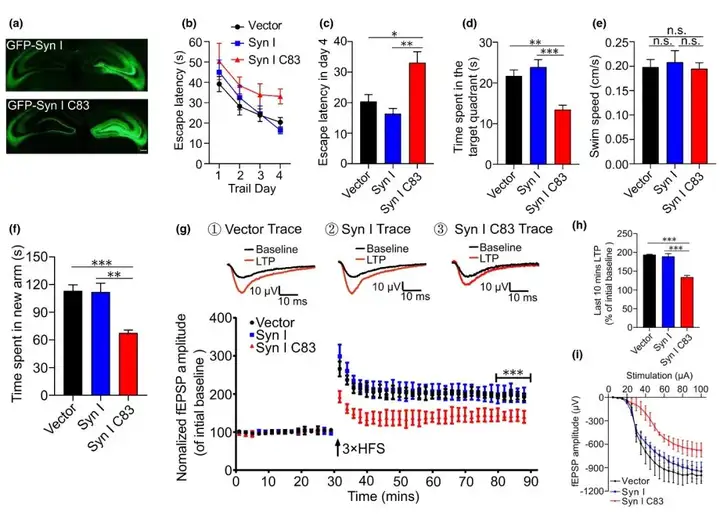
作者进一步研究AEP产生的突触素Ⅰ C83片段对tau P301S转基因小鼠(tau蛋白病小鼠模型,tau蛋白与AD相关,AD患者脑内tau蛋白异常过度磷酸化)突触功能障碍和认知损害的影响。该模型小鼠表达人突变体tau P301S,并在大脑中形成广泛的神经原纤维缠结样内含物,伴有突触功能障碍和行为障碍。将编码EGFP、EGFP-突触素Ⅰ和EGFP-突触素Ⅰ C83的AAV分别注射到2月龄tau P301S小鼠海马区域。三个月后,在所有小鼠的海马区观察到强烈的绿色荧光信号。通过Morris水迷宫实验评估突触素Ⅰ C83片段过表达对tau P301S转基因小鼠空间记忆的影响。在训练阶段(定位航行试验),表达EGFP的tau P301S转基因小鼠(即所有小鼠)找到平台的潜伏期逐渐缩短。然而,表达突触素Ⅰ C83小鼠的学习能力严重受损(第四天过表达突触素Ⅰ C83片段小鼠找到平台所需时间显著长于对照组及表达全长突触素Ⅰ小鼠)。空间探索试验中,表达突触素Ⅰ C83的小鼠出现在目标象限的时间减少,表明学习和记忆能力损伤。所有小鼠的游泳速度相当,表明全长突触素Ⅰ或其片段的过度表达不会影响运动功能。与水迷宫测试结果一致,Y迷宫测试中,表达突触素Ⅰ C83的小鼠在新异臂中花费的时间更少。海马场兴奋性突触后电位(fEPSP)的长时程增强(LTP)被认为是学习和记忆的基础。研究人员观察到表达突触素Ⅰ C83小鼠相比于表达EGFP或全长突触素Ⅰ小鼠LTP减少。与表达EGFP或全长突触素Ⅰ小鼠比较,表达突触素Ⅰ C83小鼠fEPSPs的平均振幅更低。I/O曲线显示突触素I C83小鼠的fEPSPs反应弱于其他组。这些结果表明,突触素Ⅰ C83片段的过表达诱导tau P301S转基因小鼠的突触功能障碍和认知损伤。对小鼠海马切片的电镜分析表明,表达突触素Ⅰ C83的小鼠突触密度低于表达EGFP或全长突触素Ⅰ的小鼠。此外,高尔基染色显示,突触素Ⅰ C83片段诱导AD模型小鼠的树突棘丢失。总的来说,AEP产生的突触素Ⅰ C83片段诱导tau P301S小鼠的突触功能障碍和认知障碍。
图6. 突触素Ⅰ C83片段过表达诱导tau P301S小鼠的突触功能障碍和认知功能障碍
结论
本文研究人员通过质谱分析、抗体纯化、病毒注射、行为学测试(Morris水迷宫与Y迷宫)、电生理学、电镜分析、高尔基染色等手段和方法,结合多种AD模型小鼠及AD患者标本,证明了AEP介导对Synapsin Ⅰ 的切割(产生突触素Ⅰ C83片段)诱导AD小鼠的突触功能障碍,并加速AD小鼠的病理进程。本研究揭示了AD突触功能障碍的分子机制,抑制AEP可能是一种新的、有前途的AD治疗干预措施。
武汉大学人民医院神经内科张振涛主任医师为独立通讯作者,孟兰霞博士为第一作者。本研究得到国家自然科学基金(81901090、81822016和81771382)和国家重点研发计划项目经费支持。
武汉大学人民医院张振涛教授团队合影(照片提供自:武汉大学人民医院张振涛教授团队)
通讯作者介绍
张振涛,教授,武汉大学人民医院神经内科主任医师,博士生导师,武汉大学珞珈特聘教授。国家“有突出贡献中青年专家”,“百千万人才工程”国家.级人选,享受国务院政府特殊津贴。张振涛教授课题组主要从事神经变性疾病特别是帕金森病和阿尔茨海默病的发病机制研究。该课题组主要研究领域包括:(1)天冬酰胺内肽酶(AEP)在阿尔茨海默病发病中的作用机制;(2)致病性蛋白(Tau、α-syn)在帕金森病和阿尔茨海默病发病过程中的致病机制。

本文使用的来自枢密科技的病毒产品,列表如下:


扫码下载原文
阿尔茨海默病动物模型及药效评价(更多服务及详情请咨询我们!)
阿尔茨海默病(AD)是一种起病隐匿的进行性发展的神经系统退行性疾病。临床上以记忆障碍、失语等全面性痴呆表现为特征,俗称老年痴呆症,是老年人中最常见的神经退行性疾病。它会慢慢破坏人的记忆力和思维能力,最终破坏执行简单任务的能力。随着全球老龄化加剧,AD患者数量急剧增加。而截至目前,针对AD尚无有效的治愈方法,为了加强临床医学转化研究以及临床前药物筛选,我们迫切需要发展和开发相关的动物模型。AD动物模型是以实验动物模拟AD患者脑组织的病理变化及行为特征。枢密科技可为您提供包含阿尔茨海默病模型在内多种定制大小鼠模型及表型分析服务。

了解产品及服务请扫码
或添加客服微信:BrainVTA2020

市场:027-65023363 行政/人事:027-62439686 邮箱:marketing@brainvta.com 客服:18140661572(活动咨询、售后反馈等)
销售总监:张经理 18995532642 华东区:陈经理 18013970337 华南区:王经理 13100653525 华中/西区:杨经理 18186518905 华北区:张经理 18893721749
地址:中国武汉东湖高新区光谷七路128号中科开物产业园1号楼
Copyright © 武汉枢密脑科学技术有限公司. All RIGHTS RESERVED.
鄂ICP备2021009124号 DIGITAL BY VTHINK